
-
白花鬼针草(Bidens pilosa )为菊科(Compositae)鬼针草属(Bidens)一年生的草本植物,是一种高度入侵性物种[1],原产于美洲热带地区,现已在美洲和亚洲的亚热带及热带地区广泛分布。白花鬼针草以种子繁殖为主,每株可产生3 000~6 000粒种子,种子相对较大,具有较长的倒钩刺,易附着于人或动物体表实现远距离散播[2]。此外,白花鬼针草种子萌发率高,在热带地区成熟落地后即可萌发[3],植株本身对光照和水分变化具有较高的表型可塑性[4 − 5],对极端降雨引起的短暂涝害也具有很强的适应性[6],对本地植物化感抑制作用强[7],使得其大量繁殖并快速侵占生境成为华南地区入侵面积最广泛的入侵植物之一[8]。白花鬼针草容易入侵大豆(Glycine max)、花生(Arachis hypogaea)、番薯(Ipomoea batatas)等作物农田以及郁闭度不高的果园、林地和草地等,造成土壤肥力下降、作物减产,对当地的农业、林业、畜牧业以及生态环境造成巨大影响[3]。杂草作为病原菌传播的中间体,在农业生产中扮演着重要角色。研究表明,杂草能够携带和传播多种病毒、真菌和细菌病原体,例如野燕麦(Avena fatua)携带尖孢镰刀菌(Fusarium oxysporum)和轮枝镰刀菌(Fusarium verticillioides)可传播至小麦(Triticum Linn),引发根腐病[9]。白花鬼针草的广泛分布和高适应性使其易成为病原菌的“桥梁寄主”,在农田生态系统中形成病害传播的潜在风险。白花鬼针草不仅通过竞争养分和空间影响作物生长,还能作为多种病原菌的寄主或传播媒介,进一步加剧作物病害的发生[10]。在杂草防治中,除了关注其对土壤肥力和作物产量的直接影响外,还需重视其作为病原菌传播中间体的生态学作用,采取综合防治策略以阻断病害传播链。此外,杂草病原真菌对于特定杂草的防治具有较大潜力,是新型生物除草剂极具潜力的候选资源。然而,对白花鬼针草的病原菌的研究以及生防功能的挖掘等鲜见报道。
本研究团队在调查野外杂草病害过程中发现,位于海南省儋州市农田和耕地附近的白花鬼针草种群出现了叶斑病症状,为进一步了解其病因,采用组织分离法将引起叶斑病的病原菌从病叶中分离出来,并通过形态学和分子手段对该病原菌进行鉴定,使用离体叶片接种法测定菌株的致病性,验证致病能力,研究生物学特性,旨在为白花鬼针草叶斑病菌的致病机制研究及其生物防治提供科学依据。
-
供试鬼针草植株于2023年8月采自海南省儋州市(19°51′N,109°49′E)农田和耕地附近野生种群,采集离体病叶带回实验室,100%湿度、常温条件下保存备用。供试真菌于鬼针草病叶上分离培养,用马铃薯葡萄糖琼脂培养基(potato dextrose agar, PDA)纯化培养后置于4 ℃冰箱贮藏备用。
PDA培养基(青岛海博生物技术有限公司产品);LB培养基(氯化钠10 g,酵母粉5 g,胰蛋白胨10 g,琼脂15 g,蒸馏水定容至1 L);LPGA培养基(酵母膏5 g,胰蛋白胨5 g,琼脂15 g,蔗糖5 g,蒸馏水定容至1 L);营养琼脂培养基(北京路桥生物科技有限公司产品);恰佩克培养基(Czapek)(磷酸氢二钾1 g,硝酸钠3 g,氯化钾0.5 g,硫酸镁0.5 g,蔗糖30 g,硫酸亚铁0.01 g,琼脂20 g,蒸馏水定容至1 L);1/2MS培养基(青岛海博生物技术有限公司产品)。
-
采用组织分离法对白花鬼针草的病叶分离病原真菌,供试植株中随机选取10片有病症的叶片,在样品的病健交界处切取约5 mm × 5 mm大小的叶片组织,用 w=1.0%的次氯酸钠表面消毒1 min,无菌蒸馏水冲洗3次,用无菌滤纸吸干表面水分,接种至直径为90 mm的PDA平板上,于28 ℃、L(光)︰D(暗)=12 h︰12 h光周期的条件下培养2~3 d[11]。待菌落的直径增至3 cm后,用接种环从菌落外围挑取长势旺盛的菌丝体,接种至新鲜制备的PDA培养基平板上,并通过3次连续纯化完成菌株分离,每次纯化间隔48 h,纯化获得的菌株命名为Fu-6,置于4 ℃和−80 ℃冰箱中保存。
根据柯赫氏法则对菌株Fu-6进行致病性验证,使用无菌打孔器制取直径5 mm的菌丝圆片,将其接种于经表面灭菌处理的健康植物叶片背面75%(v/v)乙醇消毒,无菌蒸馏水冲洗3次),同时设置接种灭菌PDA培养基块为空白对照,接种后置于28 ℃恒温培养箱中,采用饱和湿度环境(100% RH)进行持续保湿培养。待叶片出现病斑后,从发病部位重新分离病原菌,通过菌落形态比对及显微观察进行初步分类鉴定。将再分离菌株转接至PDA平板,28 ℃培养5~7 d后,记录菌落直径、菌丝形态、分生孢子形态及特征,并与原始接种菌株的特征进行一致性验证[12]。
-
待接种的健康叶片发病后(约接菌培养1周),用乙醇–冰醋酸混合液(V乙醇︰V冰醋酸=1︰1)固定24 h,然后在室温下转移到水合氯醛溶液(2.5 g·mL−1)中至少24 h,直到叶绿素完全除去[13]。用蒸馏水清洁叶片后,将脱色叶片放置到显微镜玻璃载玻片上固定以观察Fu-6菌株在鬼针草叶表面上的早期侵染过程。每个处理设置3次独立的生物学重复。记录病害的发病部位、发病初期症状(症状大小、形状和颜色等)和发病末期症状,使用Image软件计算叶片发病部位病斑面积。
-
根据真菌菌株的形态学特征(菌丝生长、色素沉着和真菌结构如分生孢子、分生孢子隔、分生孢子形状和大小等)对菌株进行形态学鉴定。将直径为5 mm的真菌菌块置于PDA平板(直径为9 cm)中心,并在28 ℃下于黑暗中培育5 d。对菌落外观和色素沉着情况进行拍照记录并测量,菌落直径为2个垂直直径的平均测量值。取PDA平板的少量成熟菌丝置于无菌载玻片上的水琼脂培养基薄层中,并在黑暗中于28 ℃培育15 d,使用安捷伦BioTek Cytation C10共聚焦显微镜观察菌丝和分生孢子的微形态特征并拍照记录,通过平均30个分生孢子的测量值来确定分生孢子的大小。
-
将菌株置于恒温培养箱(28 ℃)中进行培养,待菌丝体完全覆盖PDA培养基表面后,使用无菌刮铲采集菌丝体样本,经3次无菌蒸馏水漂洗后,转移至灭菌滤纸上吸除多余水分,将样本移到15 mL离心管后加入玻璃珠充分旋涡振荡混匀,采用CTAB法提取菌株的基因组DNA。使用rDNA-ITS通用引物ITS4(TCCTCCGCTTATTGATATGC)、ITS5(TCCGTAGGTGAACCTGCGG)和EF-1α引物TEF1-α-F(CCTTATCGCTCTGGTGAGGGGCATT)、TEF1-α-R(GCTCAGCGGCTTCCTGTAGTGGGGC)。分别扩增菌株的ITS、EF-1α基因序列,扩增完成后,通过1%(w/w)琼脂糖凝胶电泳检验扩增片段的大小和特异性,并将PCR扩增产物送至南山生物科技(中国)有限公司进行纯化和测序。
用NCBI数据库(
https://www.ncbi.nlm.nih.gov/ )的BLAST比对测序结果后,从GenBank数据库中下载相关菌株的序列。以咖啡青霉菌Penicillium coffeae NRRL 35363作为外群,采用MEGA 7.0软件,以最大似然法(Maximum likelihood,ML)对联合序列进行系统发育分析,Bootstraps法(重复1000 次)检验。 -
参照文献[14−15]的方法,沿菌株Fu-6的菌落边缘打取5 mm的菌块,将菌块接种于PDA培养基中央(1.4.2、1.4.3同),分别放在5、15、25、28、40 ℃的恒温培养箱中培养,每天观察1次,培养3 d后用垂直交叉测量法测定菌落直径并计算生长速率(1.4.2、1.4.5同)。
-
设置24 h持续光照、24 h持续黑暗、12 h/12 h光暗交替、24 h持续紫外光照射等4种不同光照条件,每个处理设置3组重复,恒温培养3 d后,测定菌落直径并计算生长速率。
-
选用PDA、营养琼脂(YYQZ)、1/2MS、LPGA、LB培养基,分别制备平板备用。将供试菌株接种至PDA平板中央,置于28 ℃恒温培养箱中避光培养。待菌落扩展至培养皿直径的3/4时,使用无菌打孔器沿菌落边缘切取菌块(直径D=5 mm),分别转接至上述5种培养基中央,28 ℃恒温培养,每组设置3次重复,每隔12 h观察1次菌落形态变化,采用垂直交叉测量法记录菌落直径,并计算平均生长速率。
-
采用浓度为1 mol·L−1的氢氧化钠和盐酸溶液调节PDA培养基的酸碱度,设置pH梯度为4.0~11.0(间隔1.0单位),共制备8组不同pH条件的培养基平板。将菌块分别接种于上述pH的PDA培养基中央,接种24 h后每间隔12 h记录菌丝生长情况。其他试验方法同1.4.1。
-
采用Czapek培养基作为基础基质,通过等碳当量的替代碳源(如葡萄糖、甘露醇)及等氮当量的替代氮源(如硝酸铵、蛋白胨)分别置换原配方中的蔗糖与硝酸钠组分,以无碳氮源的空白培养基作为对照组。将菌块分别接种于上述不同碳源和氮源培养基中央,接种24 h后每间隔12 h进行观察与记录。其他试验方法同1.4.1。供试碳源:可溶性淀粉、甘露醇、果糖、葡萄糖。供试氮源:酵母粉、蛋白胨、硫酸铵。
-
所有生物学特性指标均通过3次独立实验重复测定,数据以平均值±标准误形式呈现。采用SPSS 27.0软件进行数据分析,其中组间差异通过Duncan's多重范围检验进行评估,P < 0.05表示差异达显著水平。
-
以接种灭菌的PDA培养基块为对照,将病原菌Fu-6接至白花鬼针草健康叶片背面,于100%湿度、28 ℃恒温培养箱黑暗培养。待叶片出现病斑后,从发病部位重新分离病原菌,与接种菌株对比,结果发现,叶片病害症状与原始病叶症状相似。利用发病叶片对病原菌进行再次分离,能获得与初始菌株同源的致病真菌;观察再次分离得到的病原菌,其菌丝、菌落形态、分生孢子均与自然病叶分离所得菌株保持高度吻合;柯赫氏法则验证结果表明,分离物Fu-6为白花鬼针草叶斑病的病原菌。
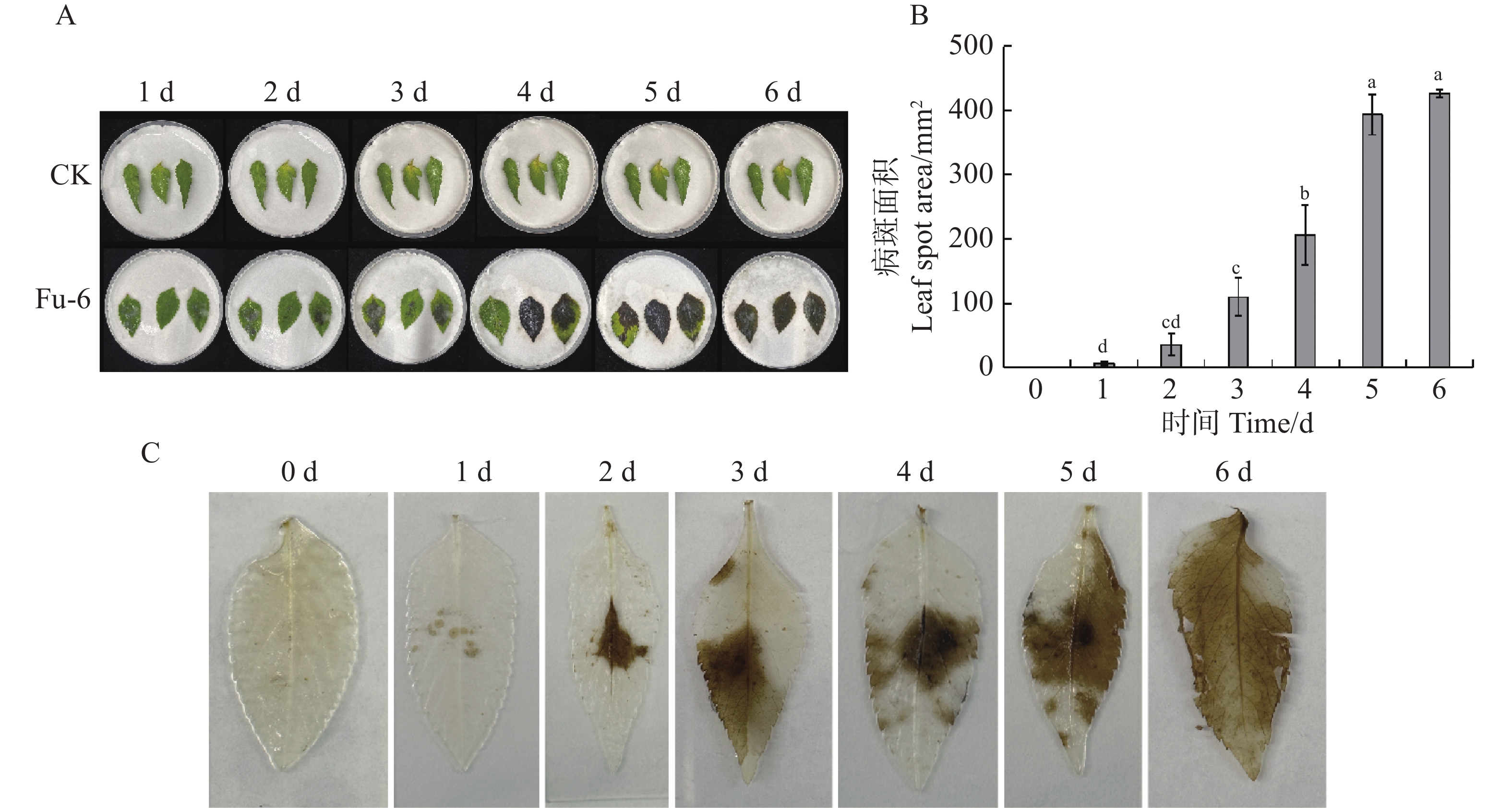
如图1所示,病原菌接种到鬼针草离体叶片2 d后,接种部位出现黄褐色椭圆病斑,呈零星点状,病斑面积为35 mm2,同时伴有白色菌丝出现;接种3 d后,病斑周边颜色加深,灰白色菌丝增多,病斑面积达109.44 mm2;4 d后,病斑由黄褐色变成黑褐色,叶片逐渐枯萎腐烂,病斑面积迅速增加;6 d后,叶片病斑面积覆盖全叶片,达到426.3 mm2(图1−A—B)。利用水合氯醛脱色法测定接种叶片的细胞死亡情况,接种1~2 d后,叶片细胞开始死亡;接种3~5 d后,发病叶片表面细胞死亡数量迅速增加;接种6 d后,叶片表面细胞大部分死亡(图1−C)。结果表明,可可毛色二孢菌株Fu-6具有较强的致病性。
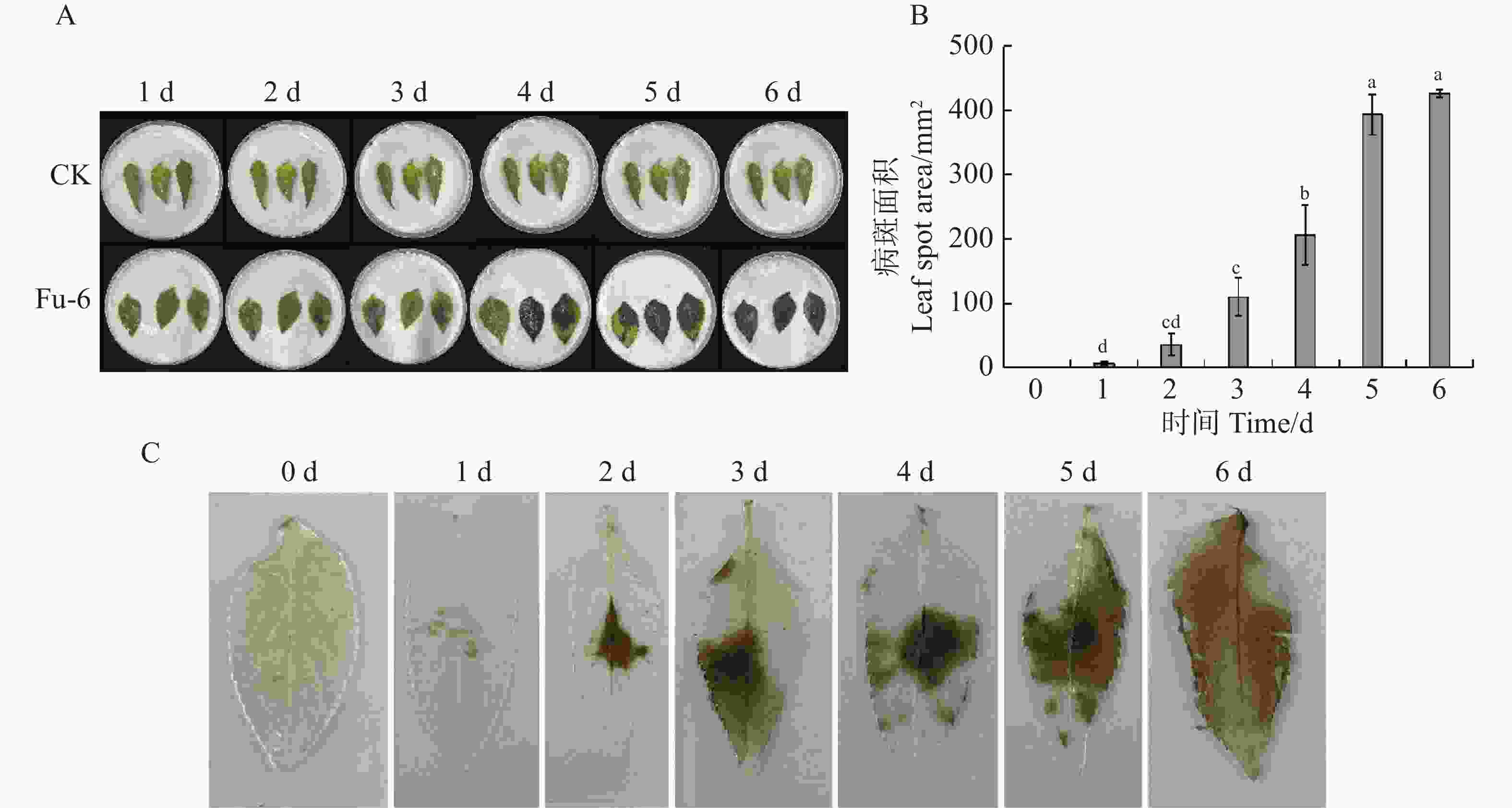
Figure 1. Determination of Bidens pilosa leaf spot pathogenicity
-
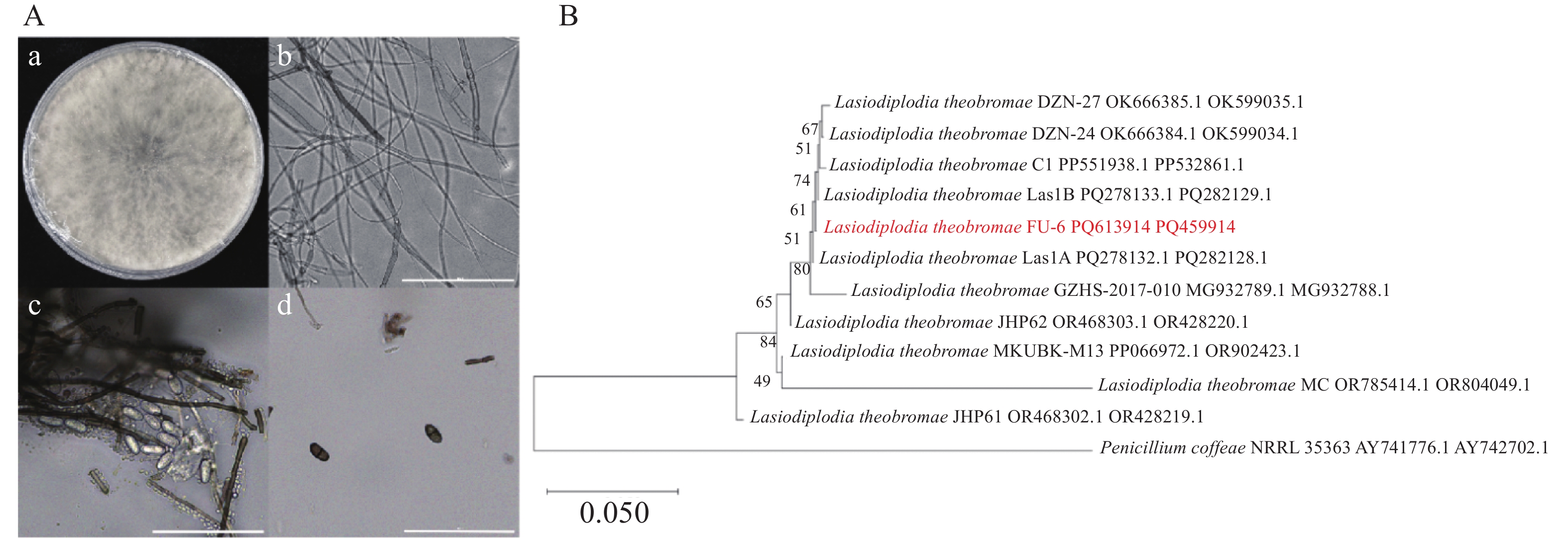
病原真菌在PDA平板生长迅速,其菌丝体在培养48~72 h即可完成对培养基表面的全覆盖(D=90 mm)。在PDA培养基上菌落为圆形,呈放射状,边缘不整齐,菌丝生长初期表面形成致密的绒毛状结构,为乳白色棉絮状菌落,气生菌丝生长旺盛,随着菌丝体成熟度增加,菌落的颜色逐渐向深褐色过渡,后期最终全部转为黑色,显微镜观察显示菌丝呈现多隔膜和分枝形态。分生孢子器呈暗黑色近球形或具不规则轮廓,半埋于基质表面。显微镜观察下,分生孢子呈椭圆形或卵形,透明单孢,未成熟孢子平均大小为(20.03 ± 0.24)μm × (10.26 ± 0.14)μm,n=30;成熟孢子平均大小为(20.58 ± 0.22)μm × (11.16 ± 0.14)μm,n=30。分生孢子在发育初期呈现半透明状态,细胞质内含有颗粒状内含物且无隔膜结构,随着成熟度增加,逐渐转变为棕褐色并形成中央横膈膜(图2−A,比例尺=100 μm)。根据菌落形态、分生孢子器形态、分生孢子规律等特征,初步判定菌株Fu-6为可可毛色二孢菌(Lasiodiplodia theobromae)。
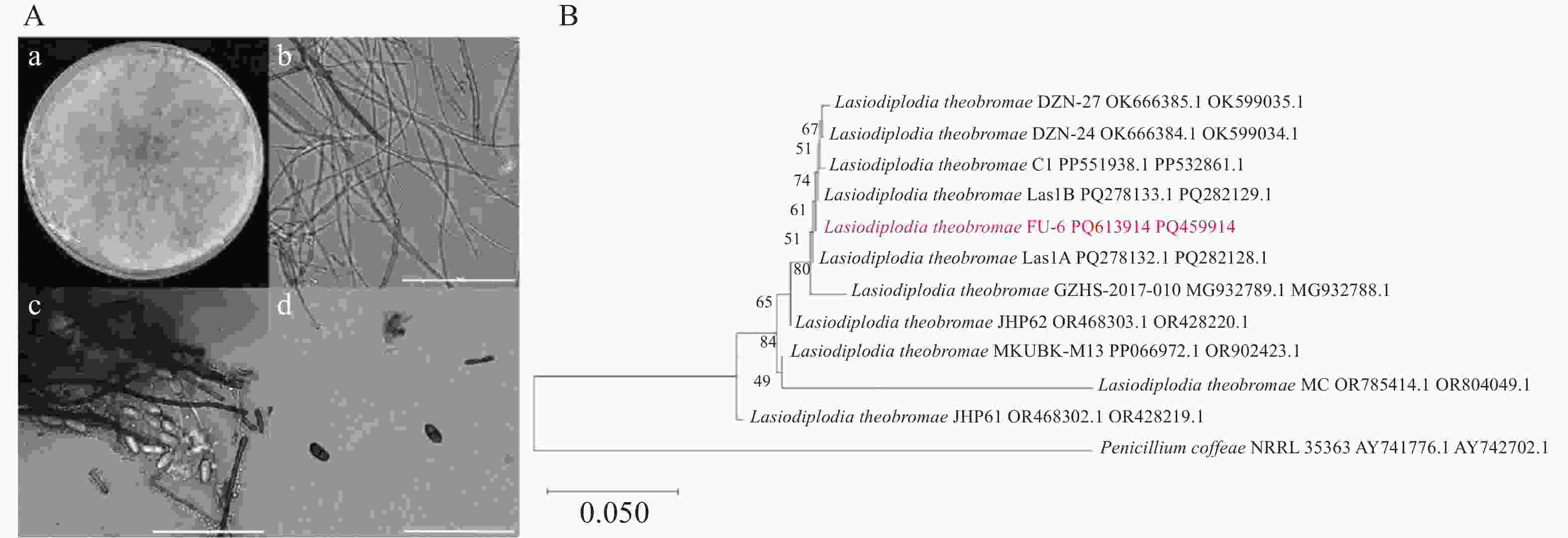
Figure 2. Morphological and molecular identification of pathogens
-
通过采用ITS4和ITS5引物进行PCR扩增,得到了长度548 bp的rDNA-ITS序列(GenBank登录号:PQ459914)。利用NCBI数据库(
https://www.ncbi.nlm.nih.gov/ )的标准核苷酸比对(BLAST)对菌株Fu-6的序列进行同源性分析,结果显示,其与可可毛色二孢(Lasiodiplodia theobromae)的参考序列存在99%的高度一致性。扩增并测序菌株tef1基因的部分序列,得到了长度172 bp的tef1序列(GenBank登录号:PP613914),将该序列与L. theobromae分离物的tef1基因的序列比对,发现相似性也为99%。选择有效发表菌株,通过ITS-tef1多序列构建系统发育树,显示Fu-6菌株与L. theobromae聚在同一分支(图2−B)。综合以上信息,Fu-6菌株被证明为可可毛色二孢菌(Lasiodiplodia theobromae)。 -
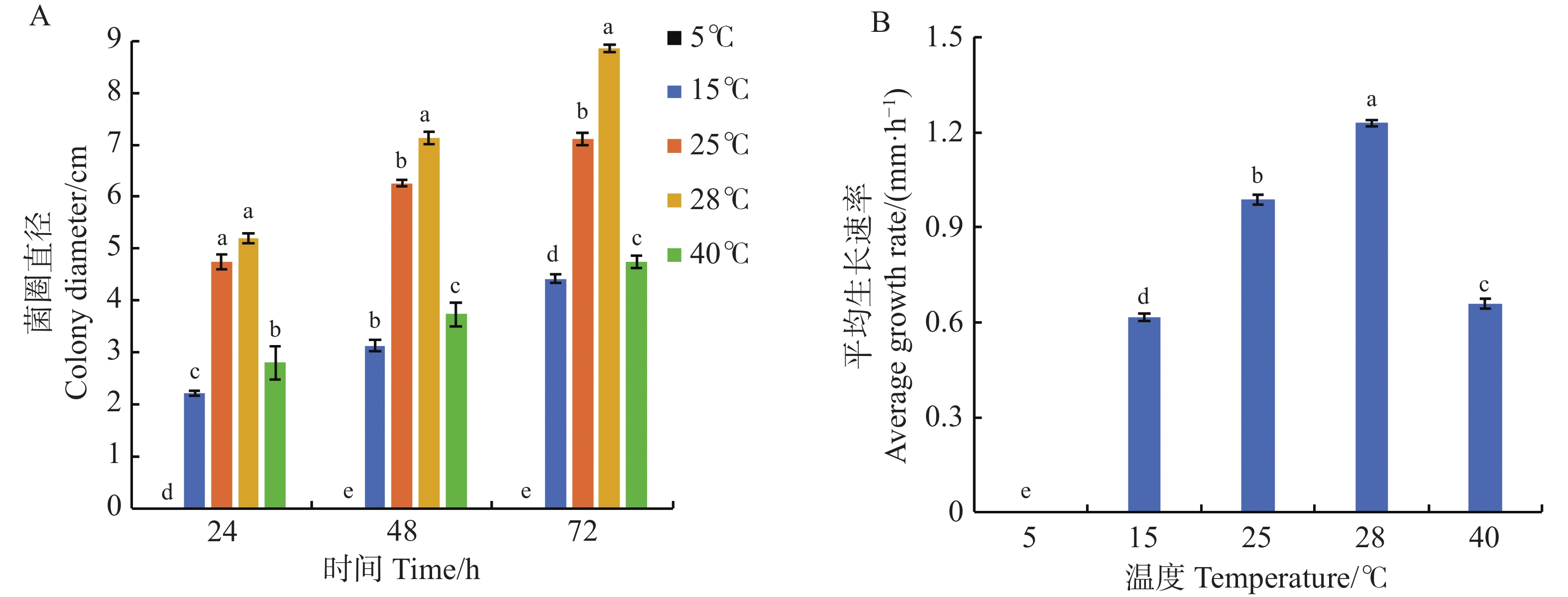
从图3可知,可可毛色二孢菌的最适生长温度为28 ℃,该温度培养时,菌株生长速率最快,为1.23 mm·h−1,72 h时菌落直径为8.85 cm;25 ℃时菌株生长速率为0.99 mm·h−1,菌落直径7.10 cm。在生长时间为24、48、72 h时,25和28 ℃的菌圈直径均显著高于5、15、40 ℃(图3−A),说明在25~28 ℃温度范围内,病原菌菌丝长势最好,生长速率最快,15 ℃和40 ℃时病原菌生长受到抑制,菌落直径仅为4.43 cm和4.75 cm,对应生长速率分别为0.61 mm·h−1和0.66 mm·h−1,5 ℃时病原菌无法生长(图3−B)。实验表明,可可毛色二孢菌的最适生长温度和最快生长速率温度均为28 ℃。
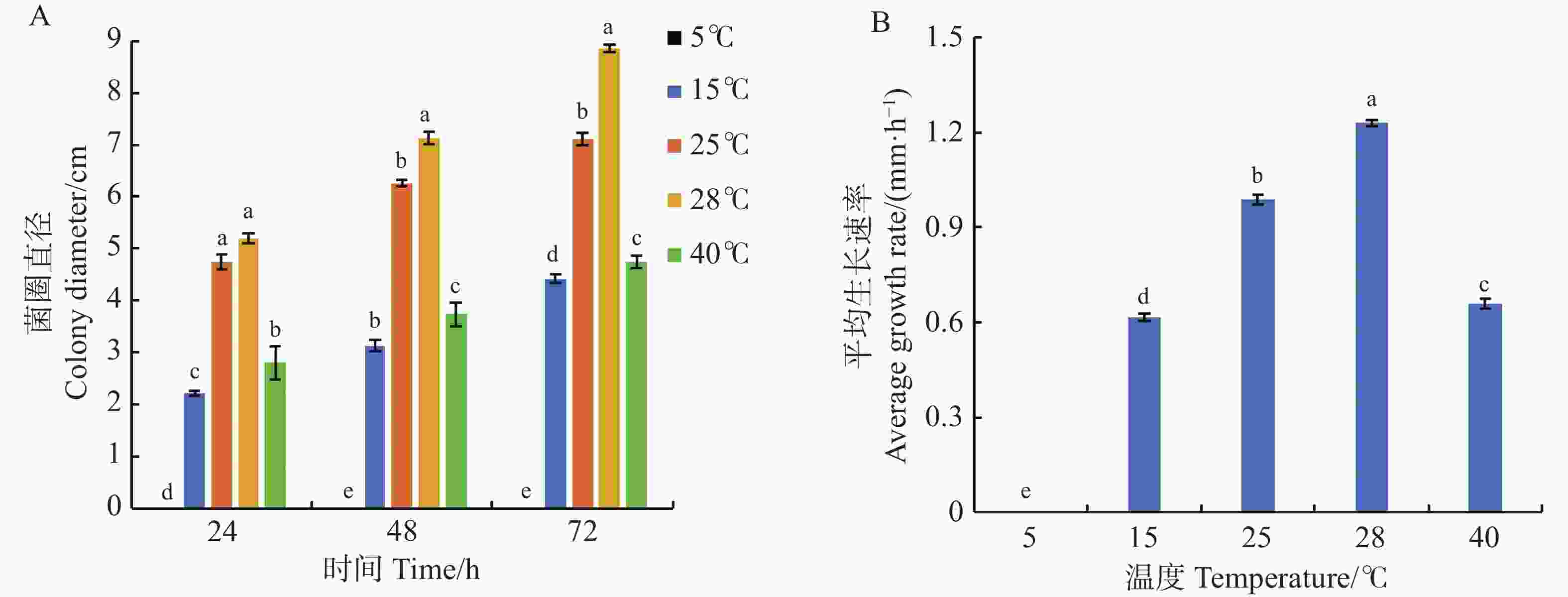
Figure 3. Effect of temperature on mycelial growth of Lasiodiplodia theobromae
-
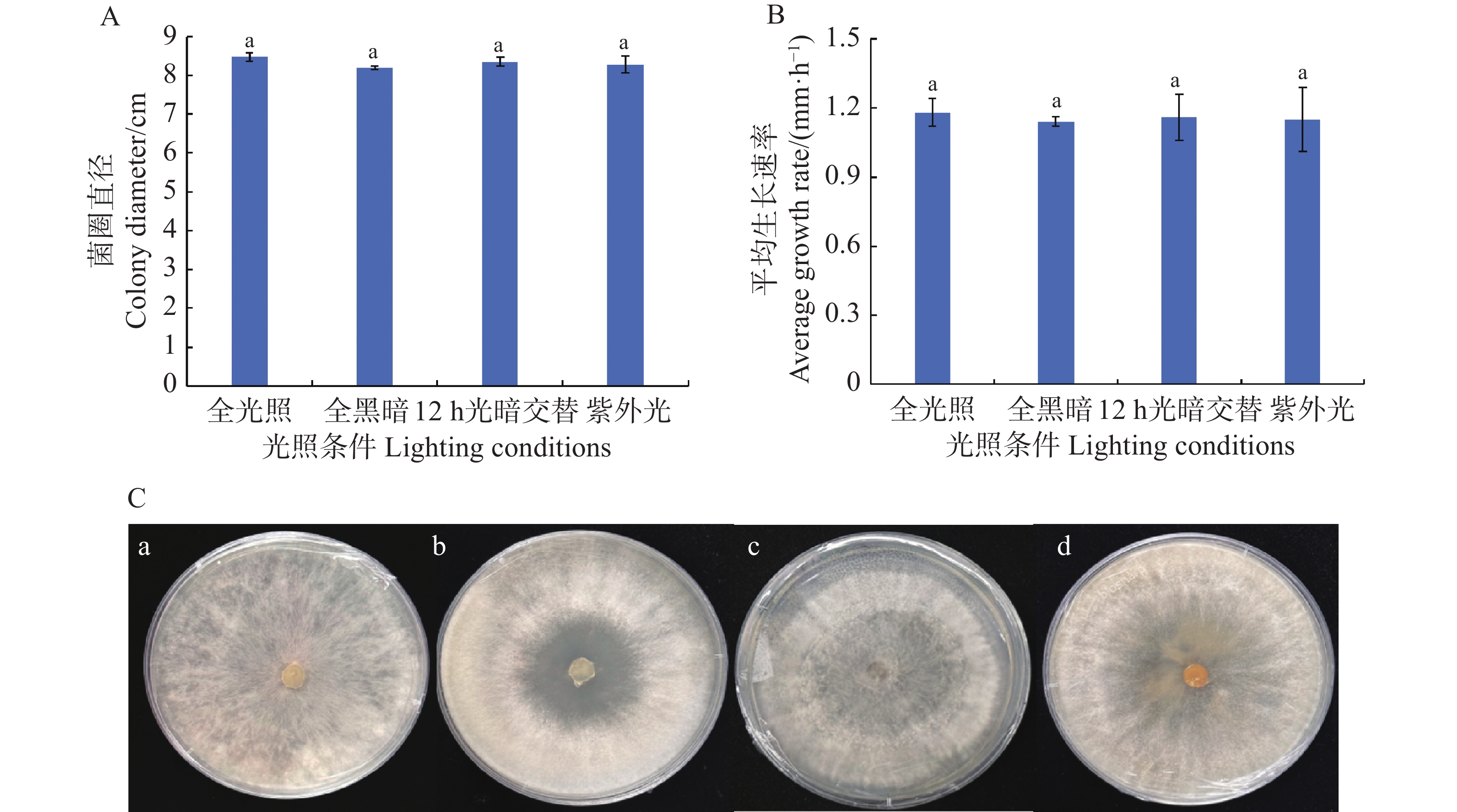
从图4可知,在连续光照(24 h·d−1)、连续黑暗(24 h·d−1)、光暗交替(12 h·d−1光照+12 h·d−1黑暗)以及持续紫外光(24 h·d−1 UV)照射等4种处理条件下,可可毛色二孢菌的菌丝均能正常生长,菌丝平均生长速率为1.14~1.18 mm·h−1,其中,连续光照生长最快(1.18 mm·h−1),连续黑暗生长最慢(1.14 mm·h−1),光暗交替和持续紫外光照射分别为1.16和1.15 mm·h−1。培养72 h后,菌丝直径分别为8.47、8.19、8.35、8.28 cm。各组间生长速率和菌落直径未出现显著性差异(P > 0.05),表明该病原菌菌丝生长对光照条件要求不严格。
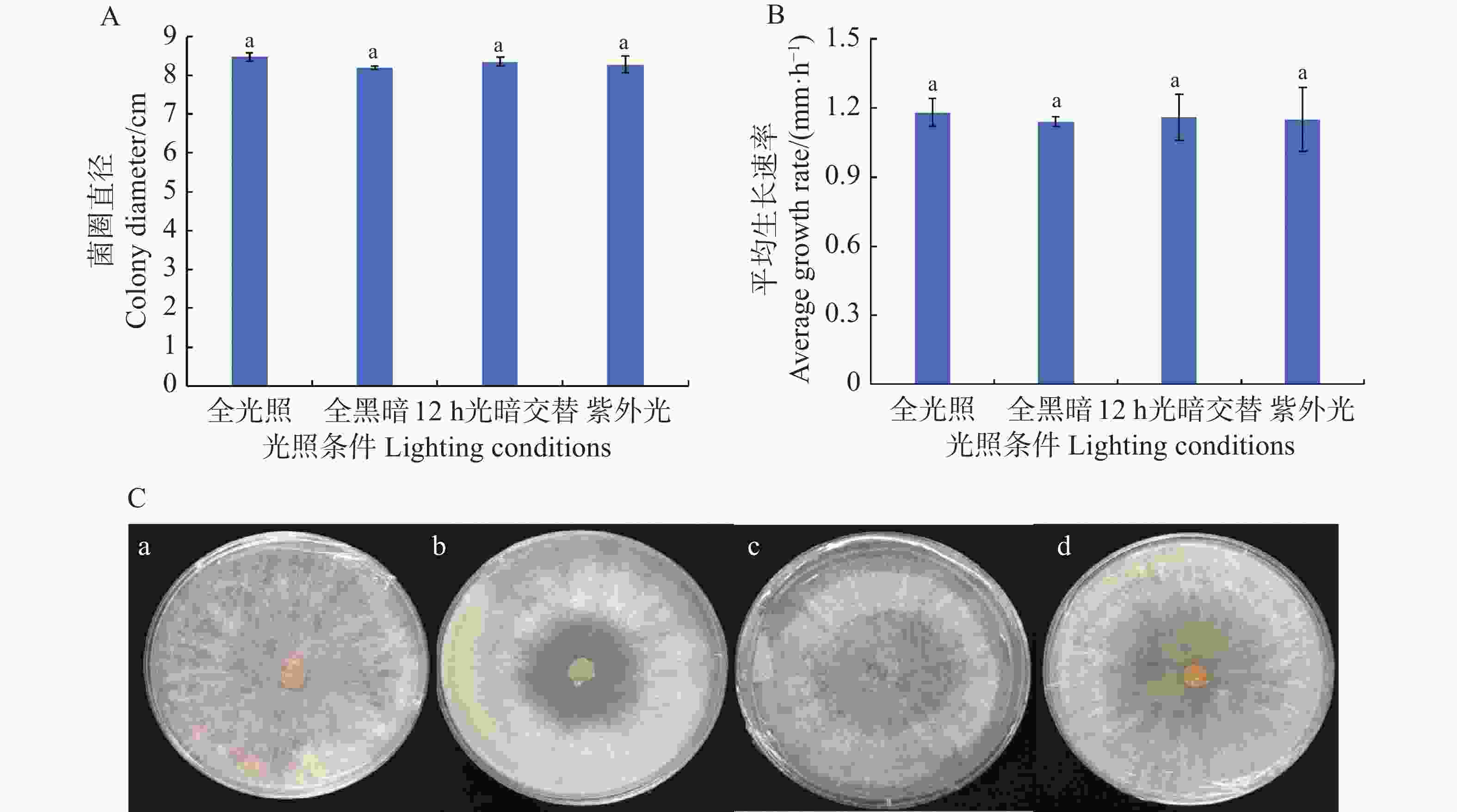
Figure 4. The effect of light on the mycelial growth of Lasiodiplodia theobromae
-
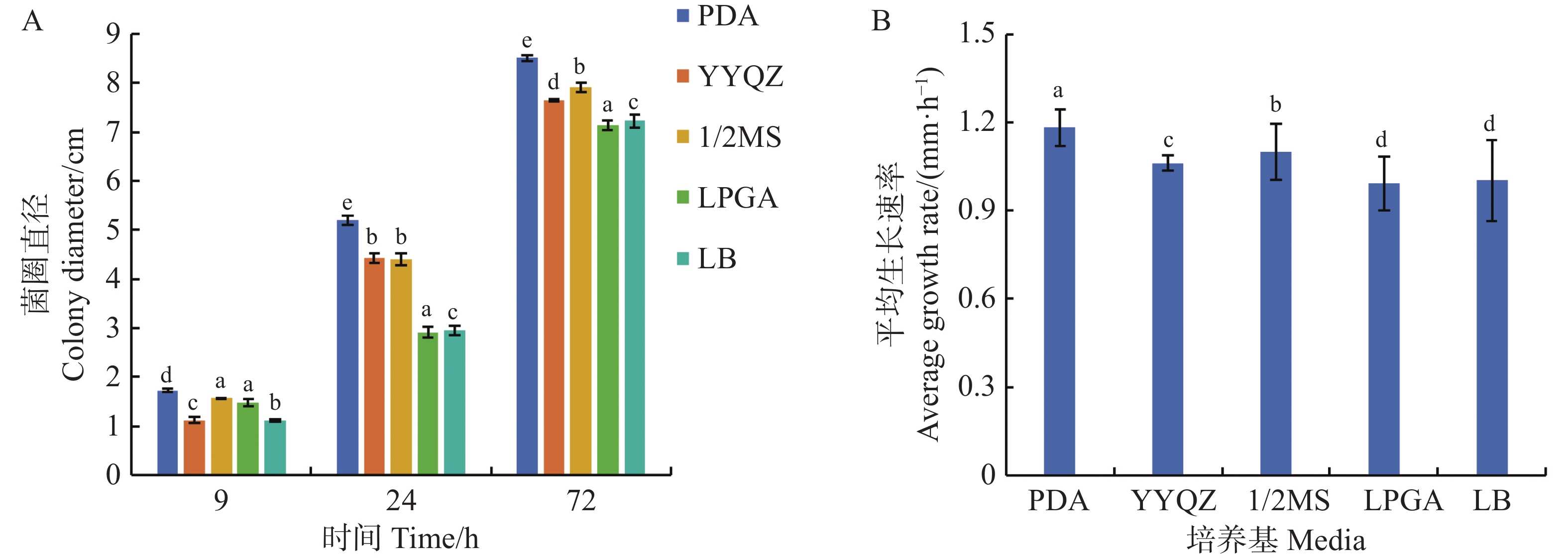
从图5可知,可可毛色二孢菌株在所有供试培养基中均能有效增殖,菌丝体在PDA培养基上扩展速率最快,平均生长速率达到1.18 mm·h−1,显著高于其他培养基,经72 h培养后菌落直径扩展至8.51 cm,其次是1/2MS培养基和营养琼脂培养基(YYQZ),菌丝生长速率分别为1.10、1.06 mm·h−1,对应的菌落直径分别为7.92、7.65 cm;在LPGA、LB的培养基中菌丝生长表现最弱,平均生长速率仅为0.99和1.00 mm·h−1,对应的菌落直径分别为7.13和7.22 cm。可可毛色二孢菌在PDA培养基上生长的3个时间点的菌圈直径和平均生长速率均高于其他4种培养基,说明马铃薯葡萄糖琼脂(PDA)培养基为菌株生长最适培养基。
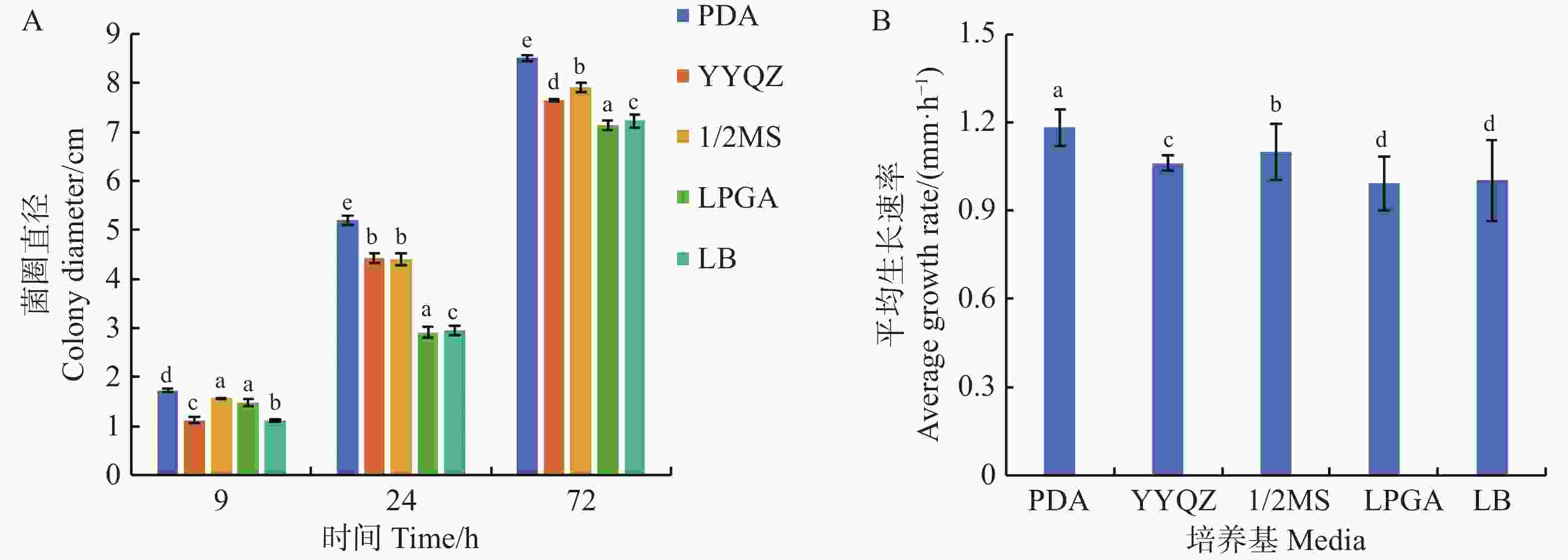
Figure 5. Effect of different media on the growth of Lasiodiplodia theobromae
-
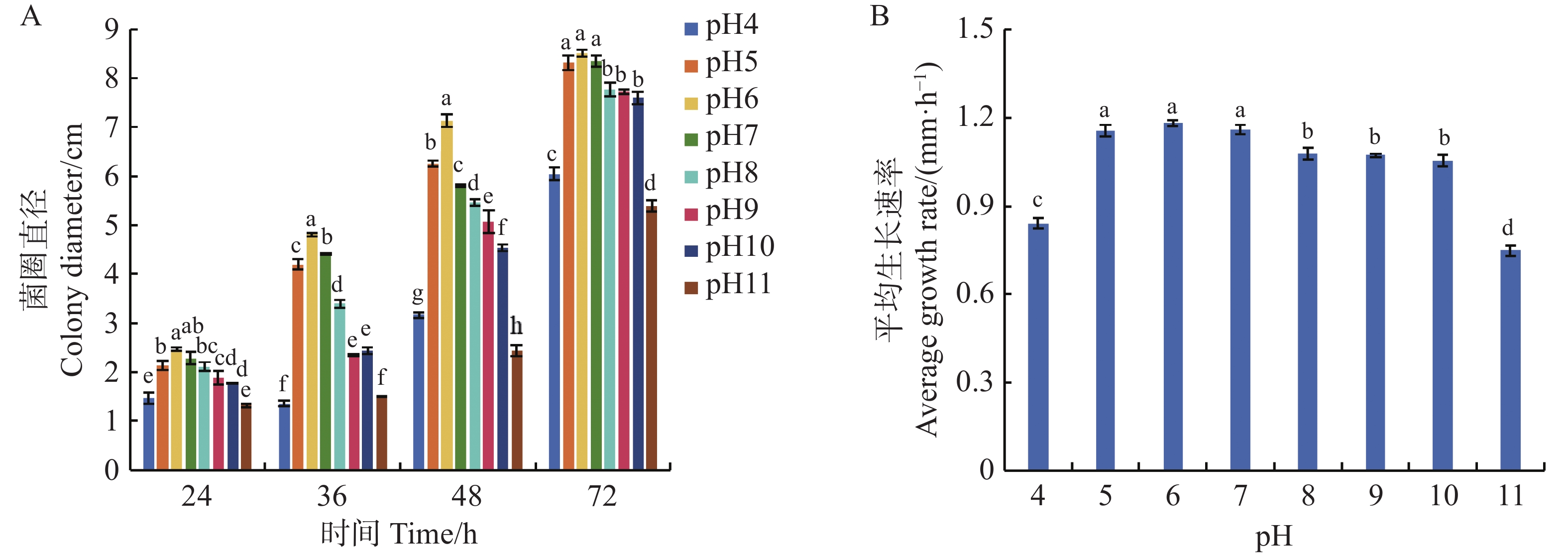
可可毛色二孢菌丝在pH4~11的PDA培养基上培养24 h均能生长,其中,在pH6的弱酸性条件下,菌丝生长速率最快,达到1.18 mm·h−1。培养48 h时菌圈直径为7.13 cm,显著优于其他pH。至72 h时,pH5~7时菌丝直径最大,菌落直径较其他处理组增加12%~18%,表明弱酸性至中性环境(pH5~7)是菌丝生长的最适区间;强酸或强碱条件(pH4、pH11)对菌丝发育产生显著抑制,平均生长速率分别降至0.84和0.75 mm·h−1,菌落直径仅为6.05和5.39 cm。结果表明,强酸性和强碱性条件不利该菌株生长(图6)。
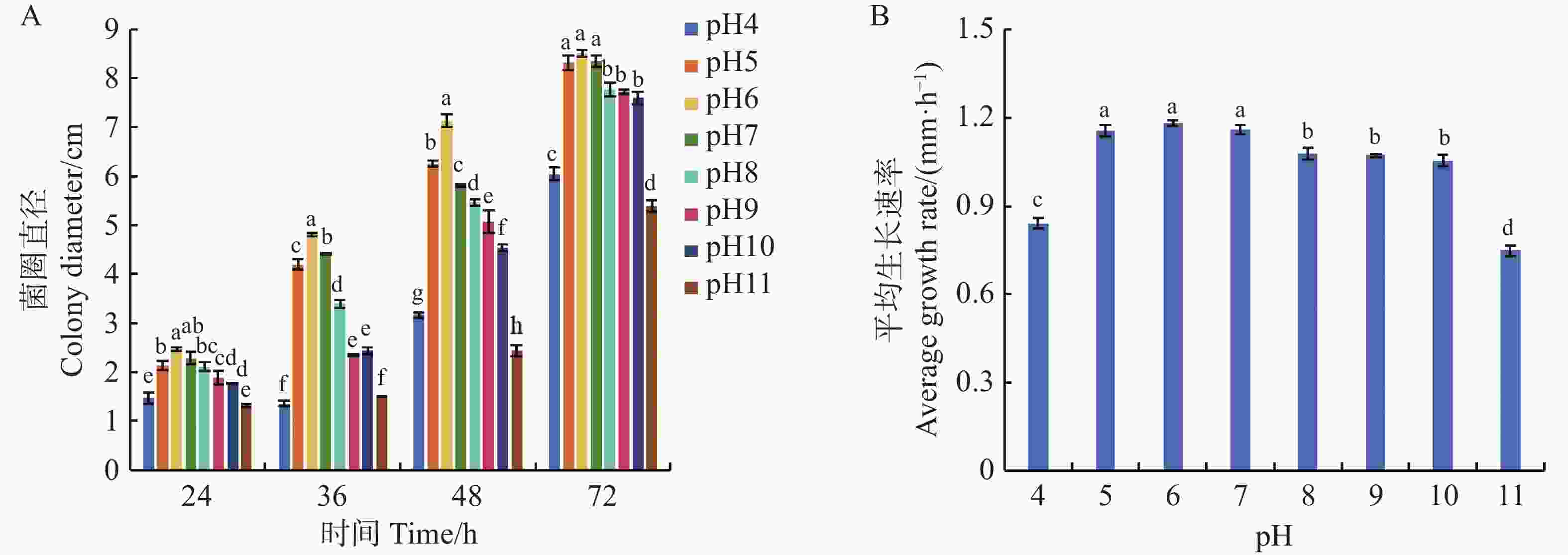
Figure 6. Effect of pH on the growth of Lasiodiplodia theobromae
-
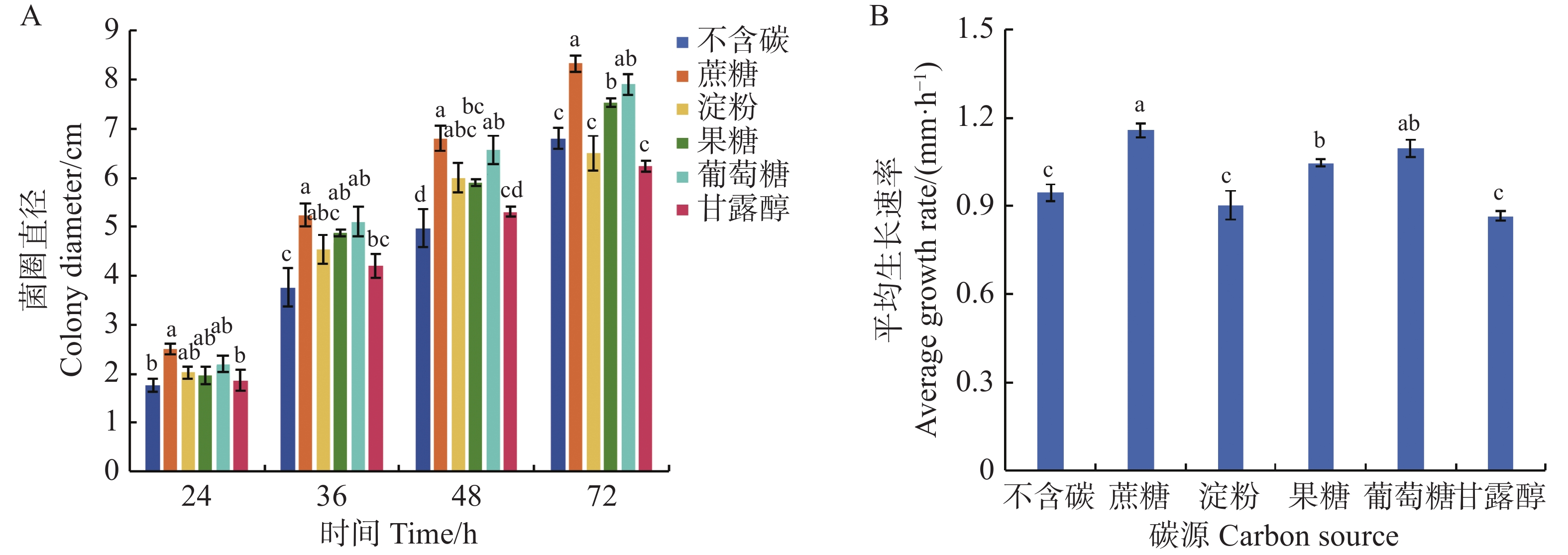
可可毛色二孢菌能利用多种碳源,其中,含糖基质对菌丝发育均呈现正向调控作用。从图7可知,在以蔗糖为碳源的培养基上菌丝生长速率达到1.16 mm·h−1,显著高于其他碳源培养基(P < 0.05),培养72 h时菌落直径为8.33 cm;其次是含葡萄糖和果糖的培养基,平均生长速率分别为1.10、1.05 mm·h−1,72 h时菌落直径为7.90、7.53 cm;在不含碳源和以淀粉、甘露醇为碳源的培养基上,菌丝平均生长速率最慢,分别为0.94、0.90、0.87 mm·h−1,72 h时菌落直径较小,分别为6.80、6.50、6.23 cm。
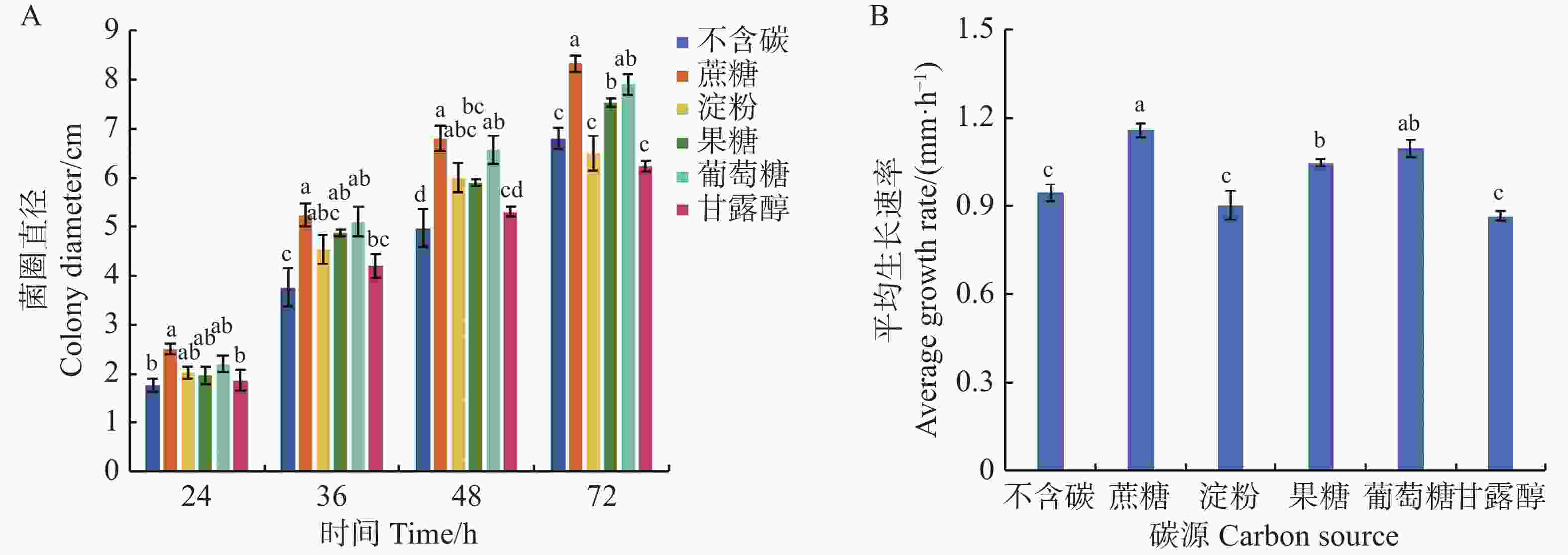
Figure 7. Effect of carbon source on mycelial growth of Lasiodiplodia theobromae
-
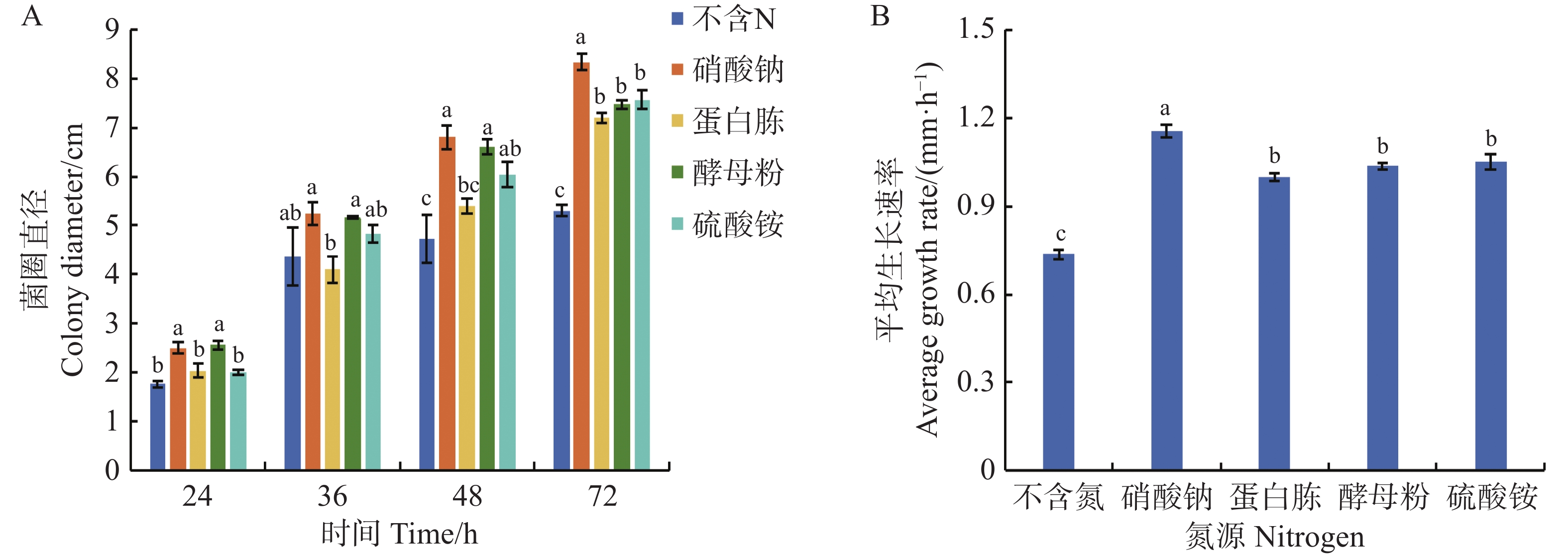
可可毛色二孢菌能利用多种氮源,实验数据显示,所有供试含氮培养基均能正向调控其菌丝发育进程。当以硝酸钠为氮源时,菌株的生长速率最快,达到了1.16 mm·h−1,培养72 h后,菌落直径扩增为8.33 cm,显著高于其他氮源培养基(P < 0.05);其次是以蛋白胨、酵母粉、硫酸铵为氮源的培养基,平均生长速率分别为1.00、1.04、1.05 mm·h−1,72 h时菌落直径为7.20、7.47、7.57 cm,无显著差异;在不含氮源的培养基上,菌丝平均生长速率最慢(0.74 mm·h−1),同时72 h时菌落直径较小,仅为5.30 cm,显著低于其他含氮源的培养基(图8)。
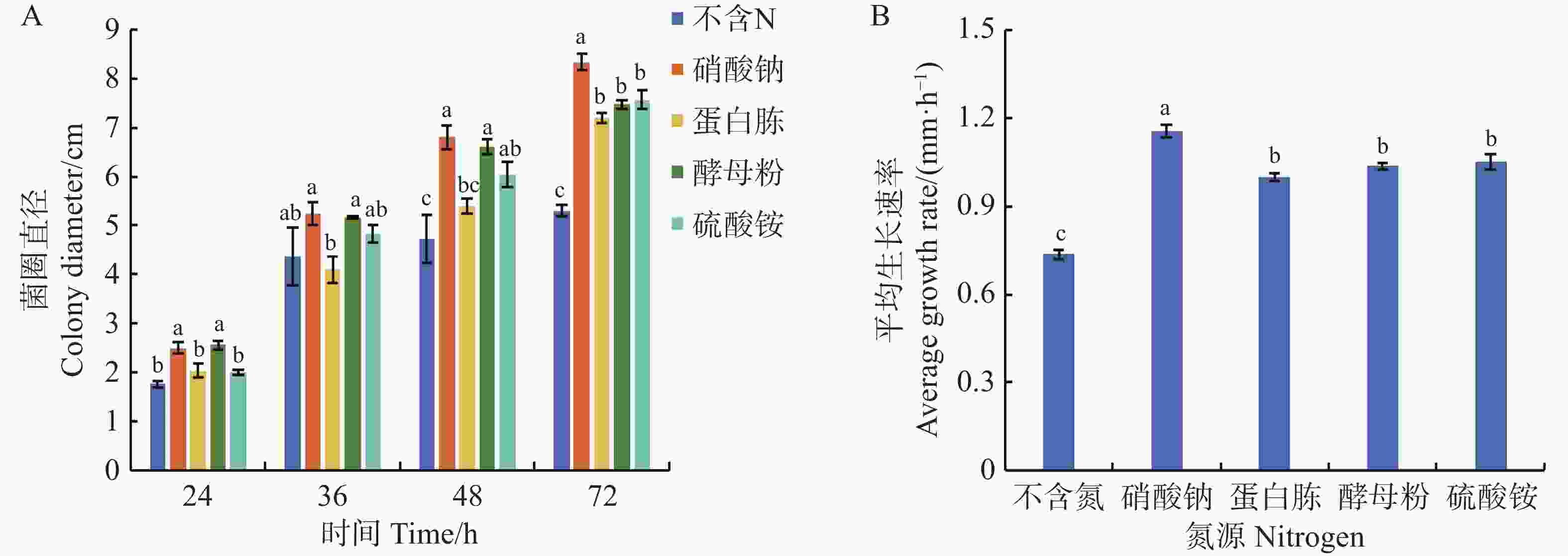
Figure 8. Effect of nitrogen source on mycelial growth of Lasiodiplodia theobromae
-
通过对引起白花鬼针草叶斑病的病原菌形态特征观察、致病性测定、分子鉴定,本研究确定白花鬼针草叶斑病的病原菌Fu-6为可可毛色二孢菌。可可毛色二孢菌是葡萄座腔菌科(Botryosphaeriaceae)中的一种广泛分布的植物病原真菌,不仅在白花鬼针草上表现出强致病性,在不同宿主植物上均表现出较强的致病性。目前,国内外已发现可可毛色二孢菌可引起葡萄(Vitis vinifera)溃疡病、芒果(Mangifera indica)蒂腐病、桃树(Prunus persica)流胶病、无花果(Ficus carica)枝干病等多种林木及果树病害[16 − 19],还可引起禾本科窄叶杂草龙爪茅(Dactyloctenium aegyptium)及菊科阔叶杂草羽芒菊(Tridax procumbens)叶片病变及枯萎[20]。可可毛色二孢菌的寄主种类多样,分布广泛,不同植物或地区分离的致病菌可可毛色二孢菌所需最适生长条件存在差异,这可能是寄主和生存的环境不同所导致的。张玉音等[21]的研究结果表明,可可毛色二孢菌JF1-5最适生长温度为25 ℃,最适培养基是PDA,最适pH6,最适的氮源和碳源分别为牛肉膏、葡萄糖。石金巧等[22]发现,可可毛色二孢菌YP-2-1的最适生长温度为28 ℃,最适pH值为5,最适光照条件为连续光照,最适的碳源和氮源分别为可溶性淀粉和酵母膏。黄艳花等[23]在广西百香果茎基腐病菌分离得到了可可毛色二孢菌32-WUB3,其菌丝生长的适宜温度为28~30 ℃,pH3~8,光照为全光照,培养基为PDA,最适的碳源和氮源分别为葡萄糖和酵母粉。本研究通过生物学特性分析明确了Fu-6的最适生长条件,其中温度为28 ℃,最适培养基为PDA,最适pH6,有利于其生长的碳源为蔗糖、氮源为硝酸钠。马晨耀[14]研究发现,光照对奶浆果树(Ficus carica)可可毛色二孢菌丝生长无明显影响,与本研究结果一致。多数分离出来的可可毛色二孢菌最适pH为3~6,说明该病原菌更容易在偏酸性的环境条件下传播,这也与白花鬼针草的地区分布一致。
植物病原真菌源除草剂是利用活体真菌或其代谢产物开发的生物除草剂,具有易降解、低毒性、对环境污染小等优点,是新型生物除草剂极具潜力的候选资源[24]。目前,在18个国家,已有36种真菌病原体作为典型的生物制剂用于杂草的生物防治,包括链格孢属(Alternaria)、韧革菌属(Chondrostereum)、炭疽菌属(Colletotrichum)、茎点霉属(Phoma)、柄锈菌属(Puccinia)和核盘菌属(Sclerotinia)[13]。中国在生物除草剂研究领域已建立起了较完善的生物除草剂研究技术体系,马唐–高粱附球菌(Epicoccum sorghinum)、苘麻–球状炭疽菌(Colletotrichum coccodes)、菟丝子–胶孢炭疽菌(Colletotrichum gloeosporioides)、空心莲子草–蕉斑镰刀菌(Fusarium stoveri)、牛筋草–双色平脐蠕孢菌(Bipolaris setariae)和麦瓶草–疫霉菌(Phytophthora nicotianae)等菌–草体系研究取得的进展,已经形成了一批具有自主知识产权的生物除草剂产品和技术成果[25 − 29]。本研究结果为基于可可毛色二孢菌的生物源除草剂提供了理论依据和前期基础,今后可以进一步探索该菌株在不同环境条件下的稳定性和应用效果,以期实现其在杂草防控中的广泛应用。
Pathogen identification and biological characteristics of leaf spot disease of Bidens pilosa
DOI: 10.15886/j.cnki.rdswxb.20250033
- Received Date: 2025-02-17
- Accepted Date: 2025-03-18
- Rev Recd Date: 2025-03-11
- Available Online: 2025-09-19
- Publish Date: 2025-11-12
-
Key words:
- Bidens pilosa /
- leaf spot disease /
- pathogen identification /
- Lasiodiplodia theobromae
Abstract: Bidens pilosa, an invasive weed commonly found in South China, is considered as a companion weeds in various upland cropping systems and serves as an intermediate host for a range of phytopathogens. This study reports the occurrence of leaf spot lesions in a population of B. pilosa. To investigate the cause, pathogens were isolated from the infected leaves using a tissue isolation method, and pathogenicity was confirmed through detached-leaf inoculation assays. A combination of morphological characterization and phylogenetic analysis of concatenated rDNA-ITS and tef1 sequences enabled accurate identification of the pathogen. The pathogen was identified as Lasiodiplodia theobromae, which causes leaf wilt and rot in B. pilosa. Preliminary environmental studies indicated that the isolated strain exhibited optimal colony growth at 28 ℃, pH 6, on Potato Dextrose Agar(PDA) medium supplemented with sucrose and sodium nitrate, which served as the preferred carbon and nitrogen sources, respectively.
| Citation: | YANG Zhichao, XIAO Yang, WU Yanna, WANG Honggang, LUO kai, CHEN Yinhua, ZHANG Rui. Pathogen identification and biological characteristics of leaf spot disease of Bidens pilosa[J]. Journal of Tropical Biology, 2025, 16(6): 819-828. doi: 10.15886/j.cnki.rdswxb.20250033

|
 DownLoad:
DownLoad: