
-
植物在生长发育过程中经常遭遇干旱、高盐和极端温度等环境胁迫,这些胁迫会导致细胞内活性氧(reactive oxygen species, ROS)的过量积累[1]。虽然ROS是一种重要的信号分子,能够激活抗逆基因的表达并帮助植物适应逆境,但其过量积累会引发细胞膜脂质过氧化、DNA损伤和蛋白质失活等一系列氧化应激反应[2]。为应对氧化应激,植物进化出复杂的抗氧化调控网络,其中,超氧化物歧化酶(superoxide dismutase, SOD)、抗坏血酸过氧化物酶(ascorbate peroxidase, APX)和过氧化物酶(catalase, CAT)等关键酶通过清除ROS维持细胞稳态[3]。
在抗氧化调控网络中,信号传递和基因表达的精确调控需要激酶和转录因子的协同作用。蔗糖非发酵-1相关蛋白激酶1(sucrose non-fermenting-1-related protein kinase 1, SnRK1)是调控植物代谢平衡和胁迫响应的核心分子[4]。SnRK1由催化亚基α、调节亚基β和γ组成,其中α亚基在SnRK1介导磷酸化过程中发挥了重要作用,其在植物中由KIN10(SnRK1.1)和KIN11(SnRK1.2)编码,不仅可以感知细胞的能量状态,还能通过代谢重编程调节细胞的代谢模式以适应不同的环境胁迫[5]。例如,AtKIN10通过调节脱落酸(abscisic acid,ABA)响应元件MYC2转录因子的活性参与到ABA介导的盐胁迫和缺氧胁迫反应中[6]。为了将激酶调控的信号传递至抗氧化基因组水平,转录因子在这一过程中发挥了桥梁作用。RAV(related to ABI3/VP1)家族转录因子属于AP2/ERF家族,不仅广泛参与植物生长发育的调控,还在胁迫响应中起到关键作用[7]。在辣椒中,CaRAV1参与调节植物对盐胁迫和干旱胁迫的抗性[8]。在水稻中,OsRAV2积极响应盐胁迫的诱导[9]。
木薯(Manihot esculenta Crantz)是大戟科植物,与甘薯、马铃薯并称为世界3大薯类作物。其贮藏根富含淀粉,不仅可用作生物能源和工业原料,还是热带和亚热带地区的重要粮食与能源作物[10]。然而,热带和亚热带种植区域往往伴随干旱、高温等非生物胁迫,持续的环境压力会引发氧化应激,导致木薯产量的显著下降[11]。已有研究表明,木薯中MeKIN10在氧化胁迫下通过直接磷酸化转录因子MeRAV1和MeRAV2,增强MeRAV1/2对抗氧化酶基因MeCAT6和MeCAT7的转录激活能力,显著提高抗氧化酶的表达水平,从而清除过量的ROS,有效缓解氧化胁迫对细胞的损害[12]。
尽管已有研究证实MeKIN10通过与MeRAV1/2互作调控其功能,但MeKIN10和MeRAV1/2的蛋白互作区域尚不清楚。因此,本研究通过对MeKIN10蛋白结构域分析、AlphaFold3互作预测及酵母双杂交实验,系统鉴定MeKIN10与MeRAV1/2的互作结构域,为深入解析木薯抗氧化调控网络提供新见解,并为培育抗氧化胁迫耐受性木薯新品种提供理论依据。
-
植物材料:本研究材料选用种植于海南大学海甸校区热带农业试验基地的木薯品种‘SC124’(South China 124)。
菌种:大肠杆菌感受态细胞DH5α、AH109酵母菌株。
载体与质粒:本实验室保存的酵母双杂交载体pGBKT7、pGADT7与质粒pGBKT7-p53、pGBKT7-lam。
试剂耗材:β-巯基还原剂、十六烷基三甲基溴化铵(CTAB)、氯化锂(LiCl)、Hifair® AdvanceFast 1st Strand cDNA Synthesis Kit反转录试剂盒(翌圣),Phusion™高保真DNA聚合酶、通用DNA纯化回收试剂盒(TIANGEN)、质粒小提试剂盒(TIANGEN)、同源重组连接酶(诺唯赞)、限制性核酸内切酶(赛默飞)等。引物合成和测序由北京擎科生物科技股份有限公司(Beijing Tsingke Biotech Co., Ltd.)完成。
-
称取2~3 g新鲜木薯叶片,在液氮中充分研磨成粉末后,按照参考文献中的方法提取总RNA[13]。使用反转录试剂盒将提取的RNA逆转录为cDNA。依据Phytozome数据库中MeKIN10和MeRAV1/2基因的编码序列,设计扩增引物(表1)。以反转录后的cDNA为模板,采用PCR技术扩增目的条带,反应体系见表2。将扩增后的产物进行琼脂糖凝胶电泳,根据DNA Marker回收与预期大小一致的目的条带。随后,分别将MeRAV1/2片段与pGBKT7载体,MeKIN10片段与pGADT7载体进行同源重组连接。反应完成后,将产物转至DH5α感受态细胞中,并涂布于含有相应抗生素的LB固体培养基上。将培养基置于37 ℃培养12 ~16 h后,挑选单克隆进行菌落PCR验证并测序。测序结果与参考序列比对后将阳性克隆置于37 ℃过夜扩大培养,次日使用试剂盒提取质粒,并进行双酶切验证。验证无误后,将质粒与菌液置于−40 ℃保存备用。
引物名称
Primer序列(5′-3′)
SequencespGBKT7-MeRAV1-F ATCTCAGAGGAGGACCTGCATATGATGGACGGAAGCTGCATAGAT pGBKT7-MeRAV1-R CTGCAGGTCGACGGATCCCCGGGCAAAGCTCCAATTATCCTCTG pGBKT7-MeRAV2-F ATCTCAGAGGAGGACCTGCATATGATGGACGGAAGCTGCACGGAT pGBKT7-MeRAV2-R CTGCAGGTCGACGGATCCCCGGGCAAAGCACCGATTATCCTCTG pGADT7-MeKIN10-F CCATGGAGGCCAGTGAATTCATGGATGGGTCAACTCACCG pGADT7-MeKIN10-R AGCTCGAGCTCGACTAAAGGACTCGGAGTTGTG pGADT7-MeKIN10a-F CCATGGAGGCCAGTGAATTCTACCAAATTACAAACTTGGTA pGADT7-MeKIN10a-R AGCTCGAGCTCGATGGATCCAAACCACGGGTGCTGACGGAT pGADT7-MeKIN10b-F CCATGGAGGCCAGTGAATTCATTGATGAAGAAATTCTCCAA pGADT7-MeKIN10b-R AGCTCGAGCTCGATGGATCCATTGTCCAATAACAAATAGTA pGADT7-MeKIN10c-F CCATGGAGGCCAGTGAATTCAGGAAATGGGCTCTTGGACTT pGADT7-MeKIN10c-R AGCTCGAGCTCGATGGATCCGAGTTGTGCAAGAAACGCTGC Table 1. Primers and their sequences
组分
Components体积/μL
Volume/μL反应条件
Reaction conditions循环数
Cycle温度/℃
Temperature/℃时间
TimecDNA 2 1 95 3 min 正向引物
Forward primer2 34 95 30 s 反向引物
Reverse primer2 55 30 s 5×FastPfu Buffer 20 72 1 min/kb dNTPs(2.5 mmol·L−1) 8 1 72 8 min FastPfu DNA Polymerase 2 1 16 终止反应
Terminate reactionddH2O 64 Table 2. PCR reaction system and conditions
-
将诱饵载体pGBKT7-MeRAV1、pGBKT7-MeRAV2和pGBKT7空载体分别转化至自制的AH109酵母感受态中,转化后的酵母菌液均匀地涂在SD/-Trp固体培养基上,倒置于28 ℃培养箱,待培养2~3 d,酵母菌落生长后,进行PCR扩增并通过凝胶电泳验证阳性克隆。挑取阳性克隆后,将菌液用超纯水稀释,设定梯度为100、10−1、10−2、10−3,并分别吸取10 μL菌液点至SD/-Trp固体培养基上,倒置于28 ℃培养箱培养2~3 d后,观察酵母菌株的生长状况,以检测MeRAV1和MeRAV2蛋白是否对酵母生长有影响。
将诱饵载体pGBKT7-MeRAV1和pGADT7空载体、pGBKT7-MeRAV2和pGADT7空载体共转入自制的AH109酵母感受态中,转化后的酵母菌液均匀的涂在SD/-Leu/-Trp固体培养基上,同时将实验室保存的阳性对照(pGBKT7-P53+pGADT7-T)和阴性对照(pGBKT7-lam+pGADT7)培养至SD/-Leu/-Trp固体培养基上,待2~3 d酵母菌落生长后,PCR扩增并进行凝胶电泳验证,挑取阳性克隆后,用超纯水将菌液稀释,设置梯度为100、10−1、10−2、10−3,每个梯度吸取10 μL菌液点至SD/-Leu/-Trp和SD/-Ade/-His/-Leu/-Trp固体培养基上,倒置于28 ℃培养箱,2~3 d后,观察酵母菌株的生长状况。检测标准:SD/-Leu/-Trp固体培养基上有实验组酵母菌落生长,SD/-Ade/-His/-Leu/-Trp固体培养基上无实验组酵母菌落生长,则可证明MeRAV1和MeRAV2蛋白无自激活活性。若SD/-Ade/-His/-Leu/-Trp固体培养基上有酵母菌落生长,则认定MeRAV1和MeRAV2蛋白存在自激活活性。
-
使用NCBI的CD-search工具(
https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi )默认参数对MeKIN10进行功能结构域分析。使用AlphaFold 3(https://alphafoldserver.com )预测蛋白互作,并用PyMOL3.1(Schrödinger,美国)进行蛋白互作结构可视化和互作位点分析。 -
分别将诱饵载体pGBKT7-MeRAV1和pGBKT7-MeRAV2与4个猎物载体共转入自制的AH109酵母感受态中,转化后的酵母菌液均匀涂在SD/-Leu/-Trp固体培养基上,待2~3 d酵母生长出阳性克隆后,挑取酵母菌落PCR扩增后进行凝胶电泳验证,将阳性克隆、阳性对照与阴性对照分别挑至超纯水中,分别稀释梯度为 100、10−1、10−2、10−3,每个梯度吸取10 μL菌液点至SD/-Leu/-Trp和SD/-Ade/-His/-Leu/-Trp固体培养基上,倒置于28 ℃培养箱进行酵母双杂交点对点验证,2~3 d后观察酵母菌株的生长状况。
-
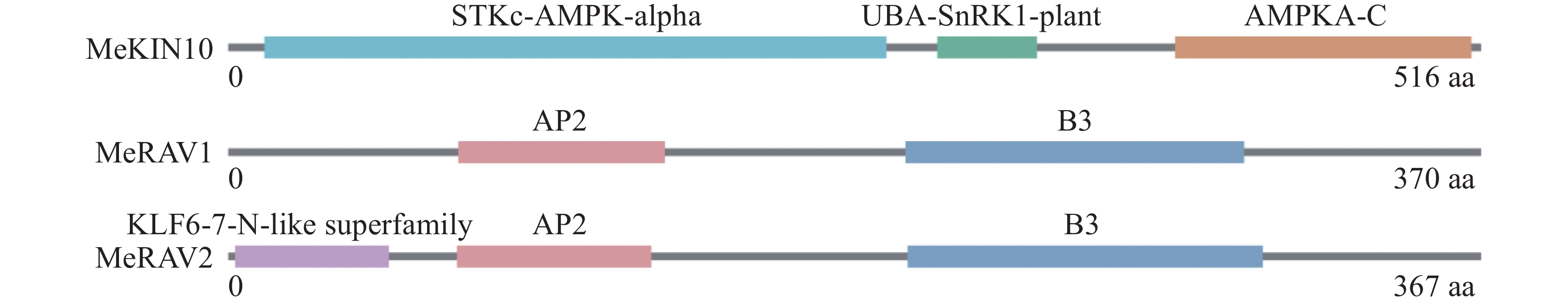
本研究使用NCBI的CD-search在线工具分析了MeKIN10和MeRAV1/2蛋白的结构域。分析结果显示,MeKIN10包含3个主要的功能性结构域。STKc-AMPK-alpha结构域是主要的激酶结构域,在调控细胞能量代谢和维持能量平衡中发挥关键作用;UBA-SnRK1-plant结构域与泛素化相关;AMPKA-C结构域在SnRK1的激酶活性调控机制中起关键作用。
MeRAV1具有两个结构域,分别为AP2和B3结构域。AP2和B3结构域为植物转录因子中常见的DNA结合域,涉及植物发育调控及对环境变化的响应。MeRAV2包含3个结构域:KLF6-7-N-like superfamily结构域(具有类似锌指的功能,参与基因表达调控),以及AP2和B3结构域(图1)。
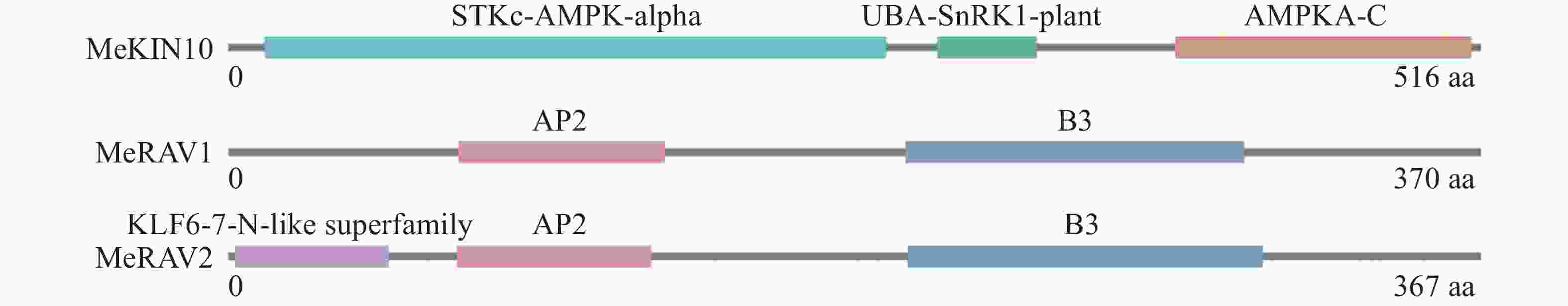
Figure 1. Protein domain analysis of MeKIN10 and MeRAV1/2
-
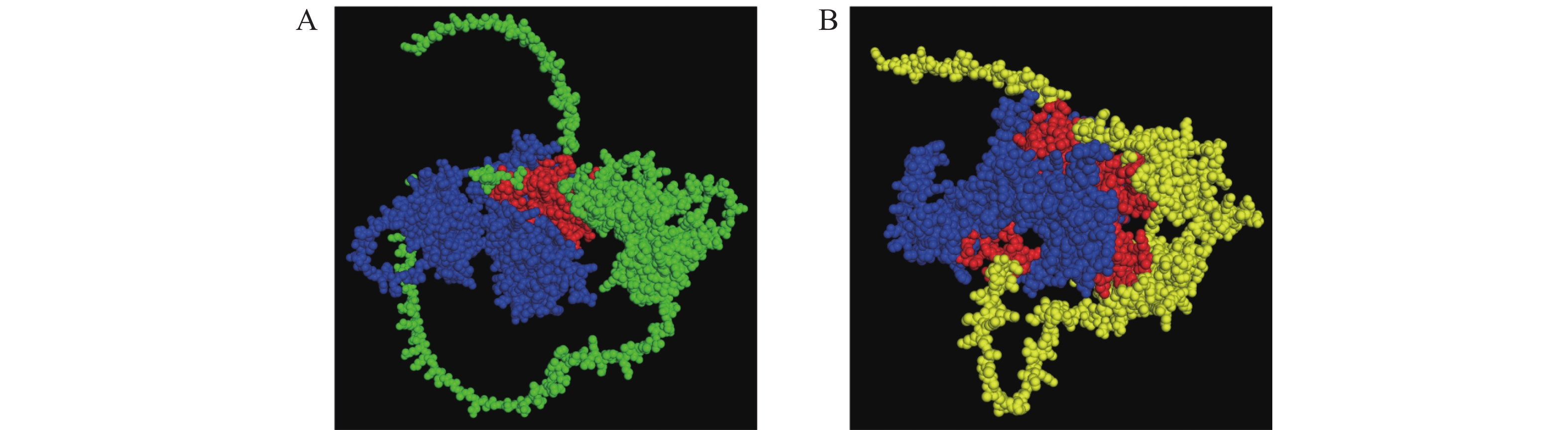
为研究MeKIN10与MeRAV1/2蛋白的相互作用位点,使用AlphaFold 3模拟其三维结构及蛋白互作。预测结果表明,MeKIN10的STKc-AMPK-alpha结构域与MeRAV1发生相互作用,而其3个结构域均与MeRAV2发生互作(图2、图3)。
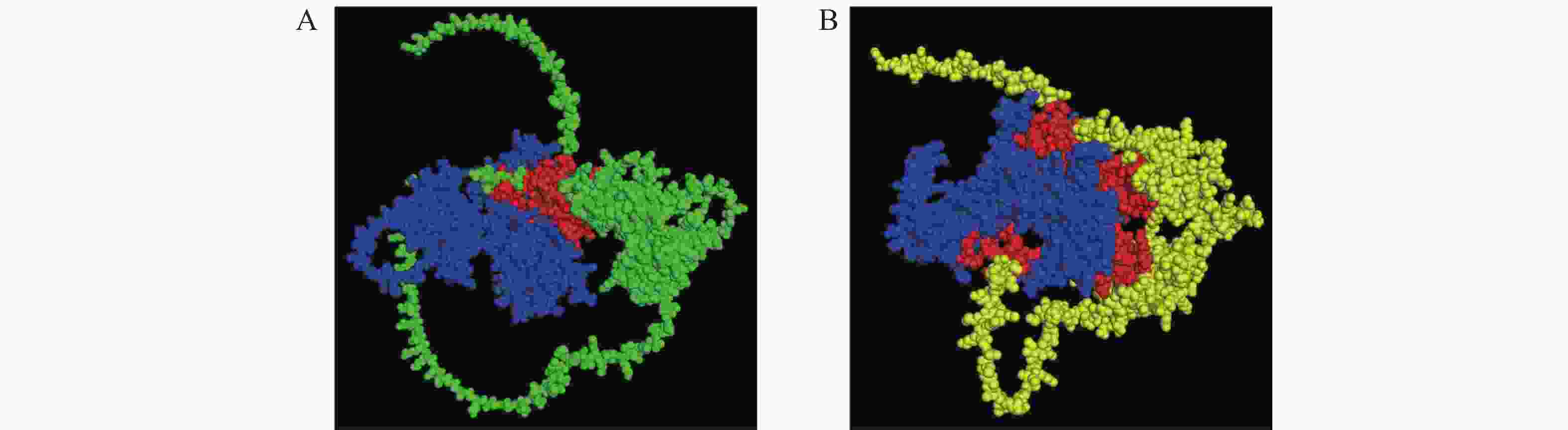
Figure 2. Three-dimensional structure and interaction prediction of MeKIN10 and MeRAV1/2 proteins

Figure 3. Analysis of the interaction sites between MeKIN10 and MeRAV1/2 proteins
-
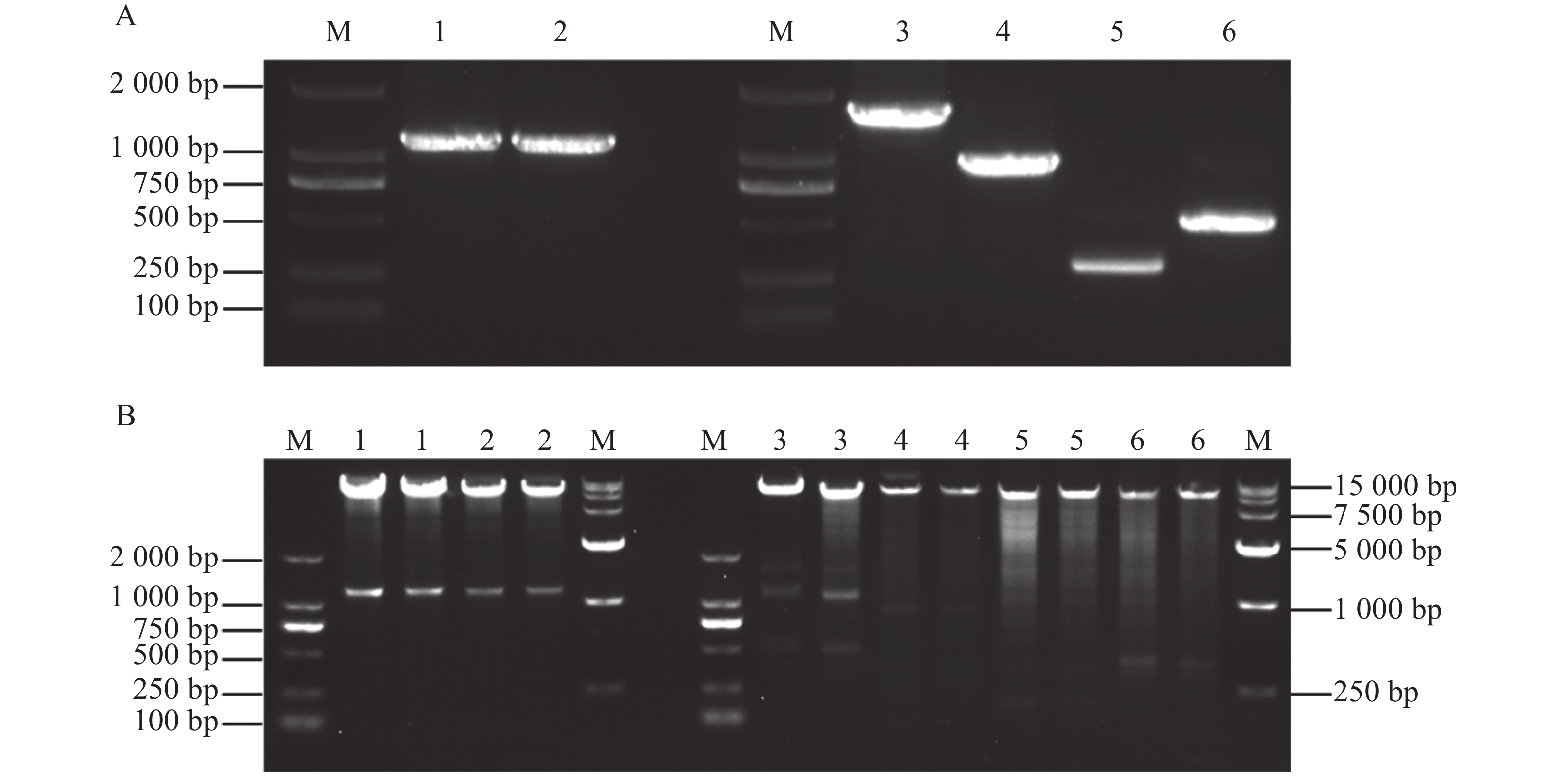
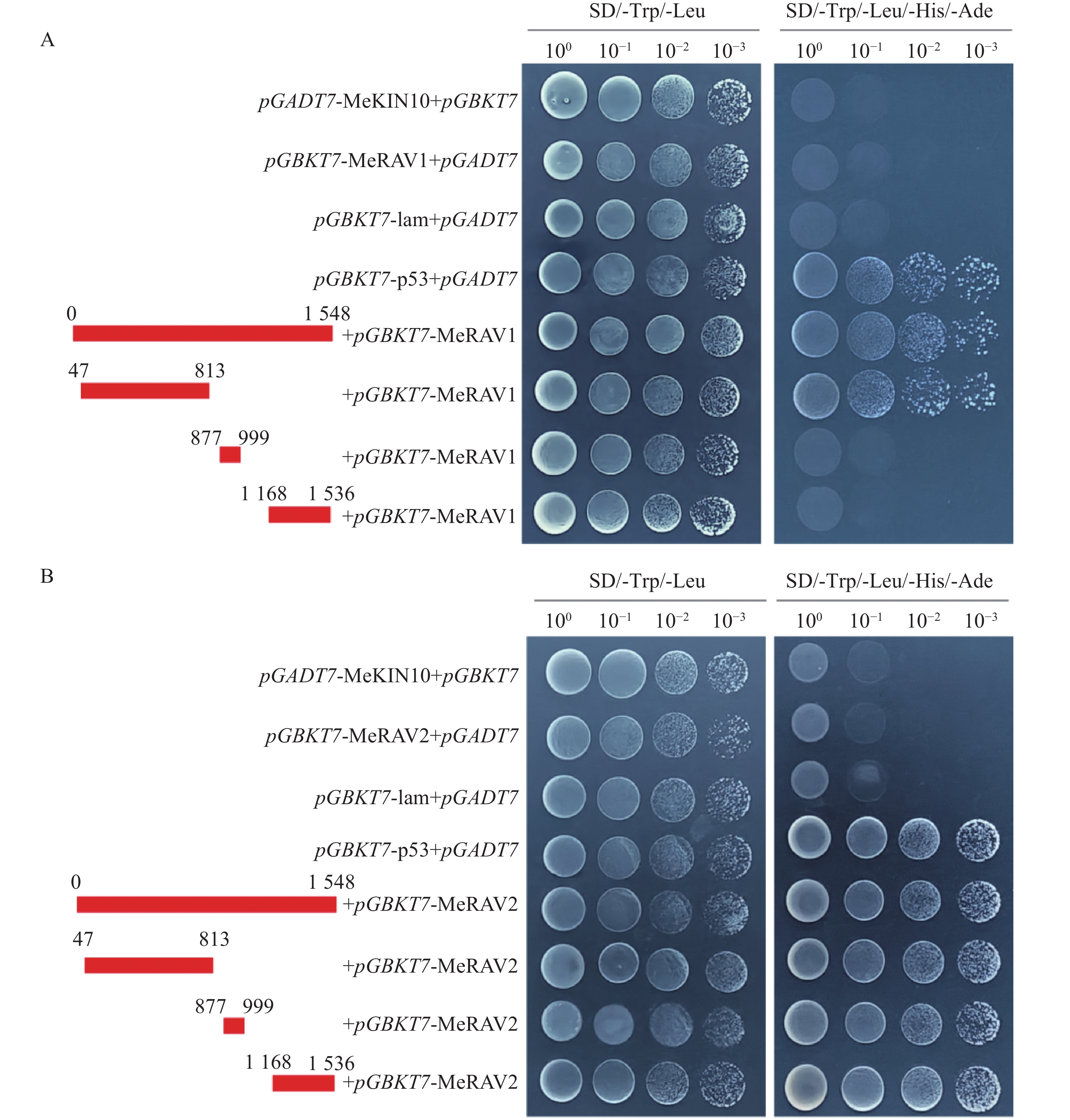
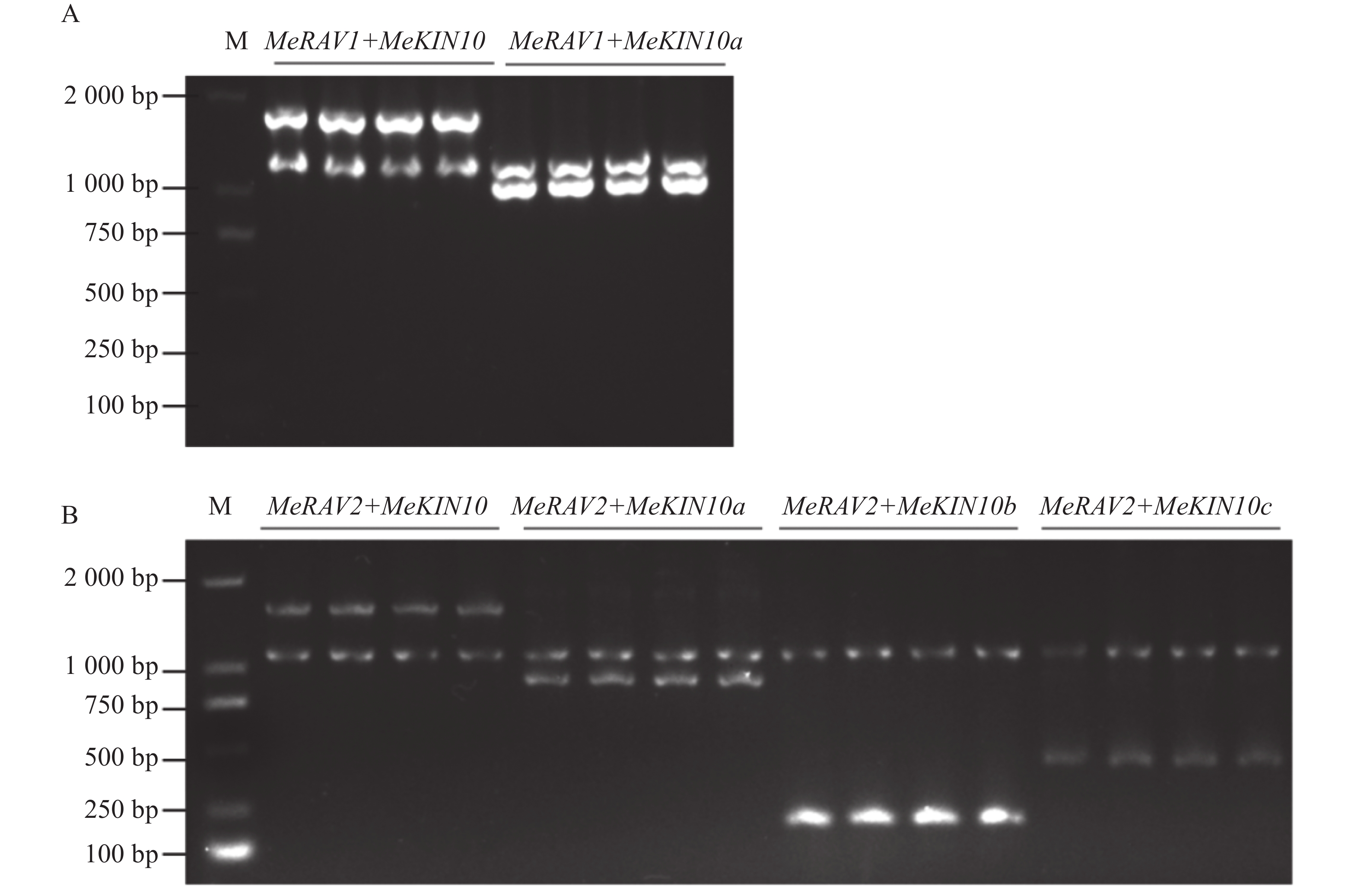
为探究MeKIN10与MeRAV1/2蛋白互作中的主要区域,基于MeKIN10蛋白的3个结构域,笔者将其分为3个基因片段,并分别构建到pGADT7载体中。得到的猎物载体分别命名为pGADT7-MeKIN10a(STKc-AMPK-alpha结构域47~813 bp)、pGADT7-MeKIN10b(UBA-SnRK1-plant结构域877~999 bp)及pGADT7-MeKIN10c(AMPKA-C结构域1 168~1 536 bp),将MeRAV1和MeRAV2的编码区构建到pGBKT7载体上,得到的诱饵载体命名为pGBKT7-MeRAV1和pGBKT7-MeRAV2。目的片段扩增结果通过琼脂糖凝胶电泳检测确认,在目标区域出现预期条带,其中MeRAV1(1 110 bp)、MeRAV2(1 101 bp)、MeKIN10(1 548 bp)、MeKIN10a(767 bp),MeKIN10b(122 bp)和MeKIN10c(368 bp)(图4−A)。使用DNA产物纯化回收试剂盒回收目的片段,后将回收的MeKIN10、MeKIN10a、MeKIN10b和MeKIN10c片段用EcoRⅠBamHⅠ双酶切好的pGADT7线性化载体进行同源重组连接;MeRAV1/2片段与用NdeⅠ和SmaⅠ双酶切好的载体pGBKT7线性化载体进行同源重组连接。将连接产物转化至DH5α感受态细胞中,放置于37 ℃下培养12~16 h后,挑选单克隆进行菌落PCR验证并测序。对测序结果正确的菌液,在37 ℃过夜扩增培养后提取质粒,最后进行双酶切验证,MeKIN10在1 066 bp位置有与EcoRⅠ相同的碱基序列,因此酶切过程中将MeKIN10切成了3段,分别是1 548、1 066、482 bp,符合预期结果,其他均切出一条符合基因片段大小的片段(图4−B)。验证无误后,选取质粒与一致菌液置于−40 ℃保存备用。
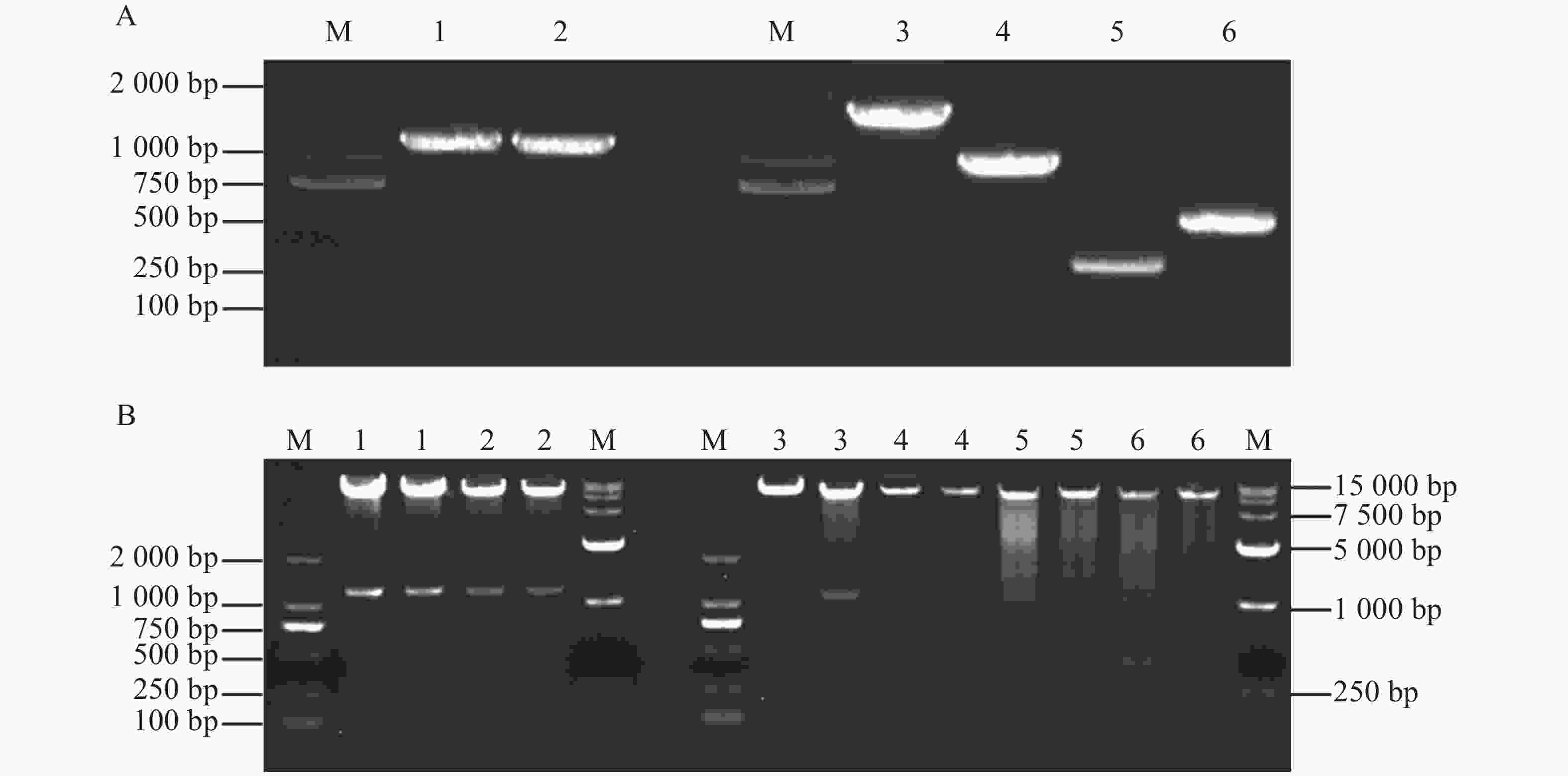
Figure 4. Construction of recombinant vector containing the target gene
-
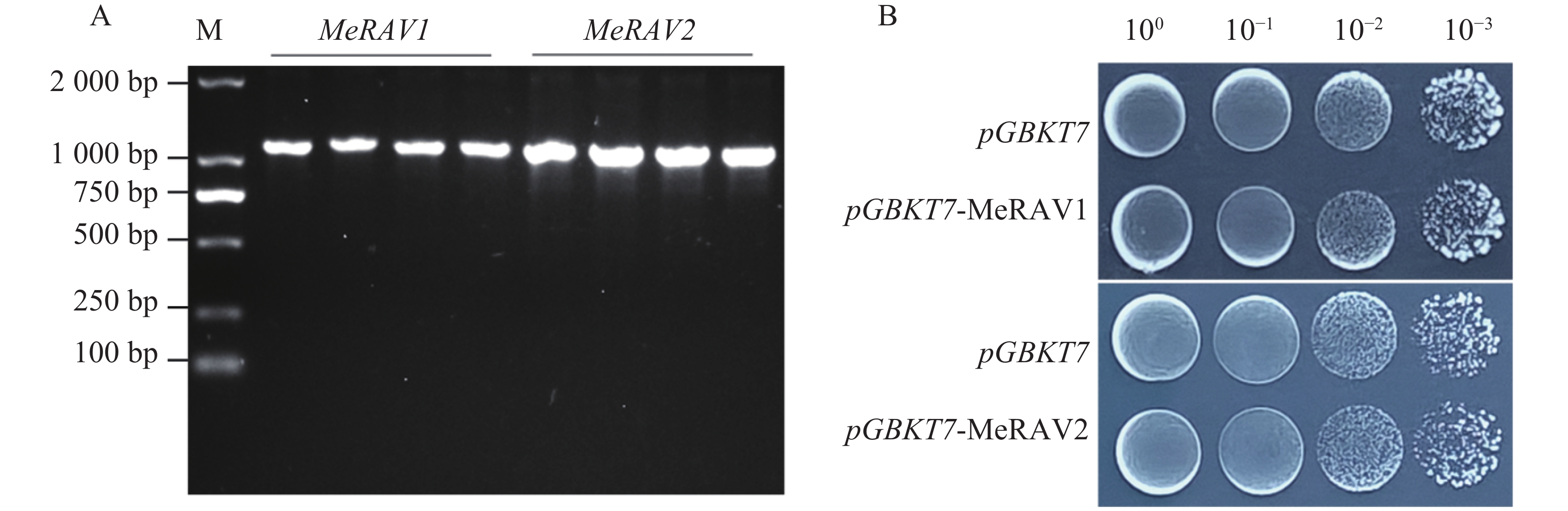
为检测诱饵蛋白是否具有毒性,将载体pGBKT7-MeRAV1/2和空载体pGBKT7分别转化至AH109酵母感受态中,通过菌落PCR筛选阳性克隆(图5−A)。将阳性克隆培养至OD600=0.8后,用超纯水稀释至10、10−1、10−2、10−3的梯度,每个梯度取10 μL菌液点滴至SD/-Trp平板。结果显示,携带诱饵载体pGBKT7-MeRAV1/2的酵母菌株与携带空载体pGBKT7的酵母菌株在生长趋势上保持一致(图5−B)。表明pGBKT7-MeRAV1/2诱饵蛋白对酵母细胞生长无显著影响,也未表现出毒害作用,表明该诱饵蛋白适合用于后续实验。
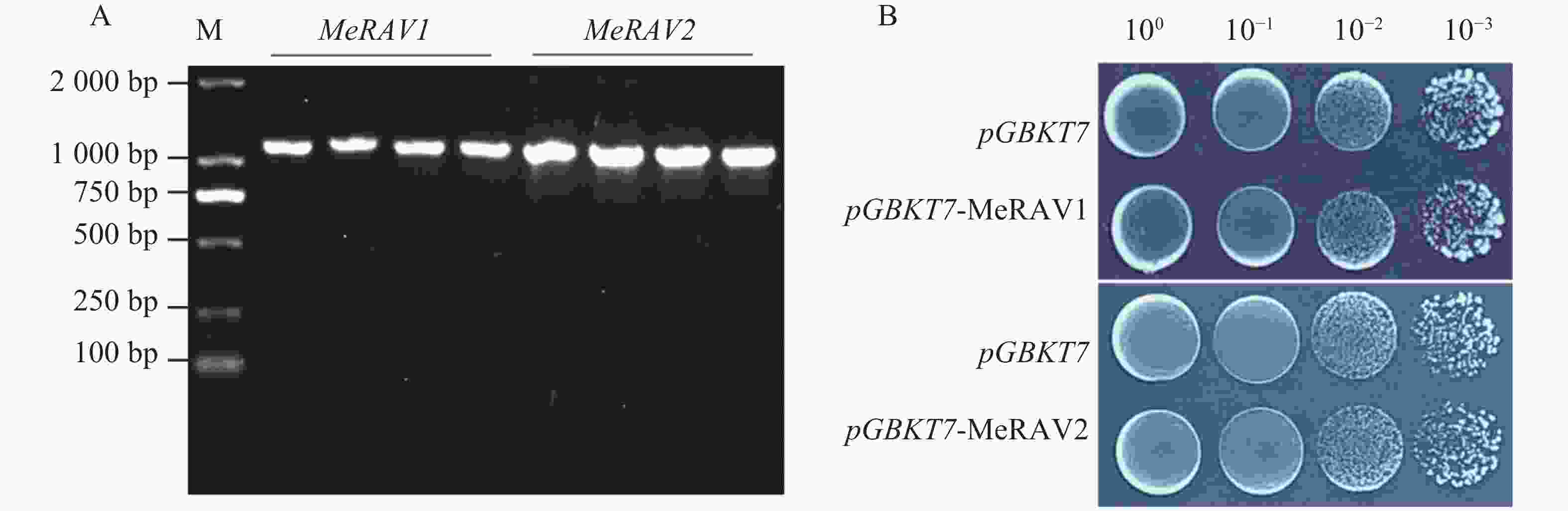
Figure 5. Toxicity test of the bait proteins MeRAV1/2
为检测诱饵蛋白是否具备转录激活活性,将pGBKT7-MeRAV1/2与空载体pGADT7共转化至AH109酵母感受态细胞中,并涂布在二缺培养基SD/-Trp/-Leu平板上,对正常生长出的酵母菌株进行PCR检测,结果显示均为阳性(图6−A),证明质粒转化成功。将阳性克隆点滴在四缺培养基SD/-Trp/-Leu/-His/-Ade平板上,结果显示,仅阳性对照(pGBKT7-p53+pGADT7)酵母菌株正常生长,阴性对照(pGBKT7-lam+pGADT7)及实验组(pGBKT7-MeRAV1/2+pGADT7)酵母菌株均未正常生长(图6−B)。这些结果表明,MeRAV1/2诱饵蛋白不具备自激活活性,适用于后续蛋白互作实验。
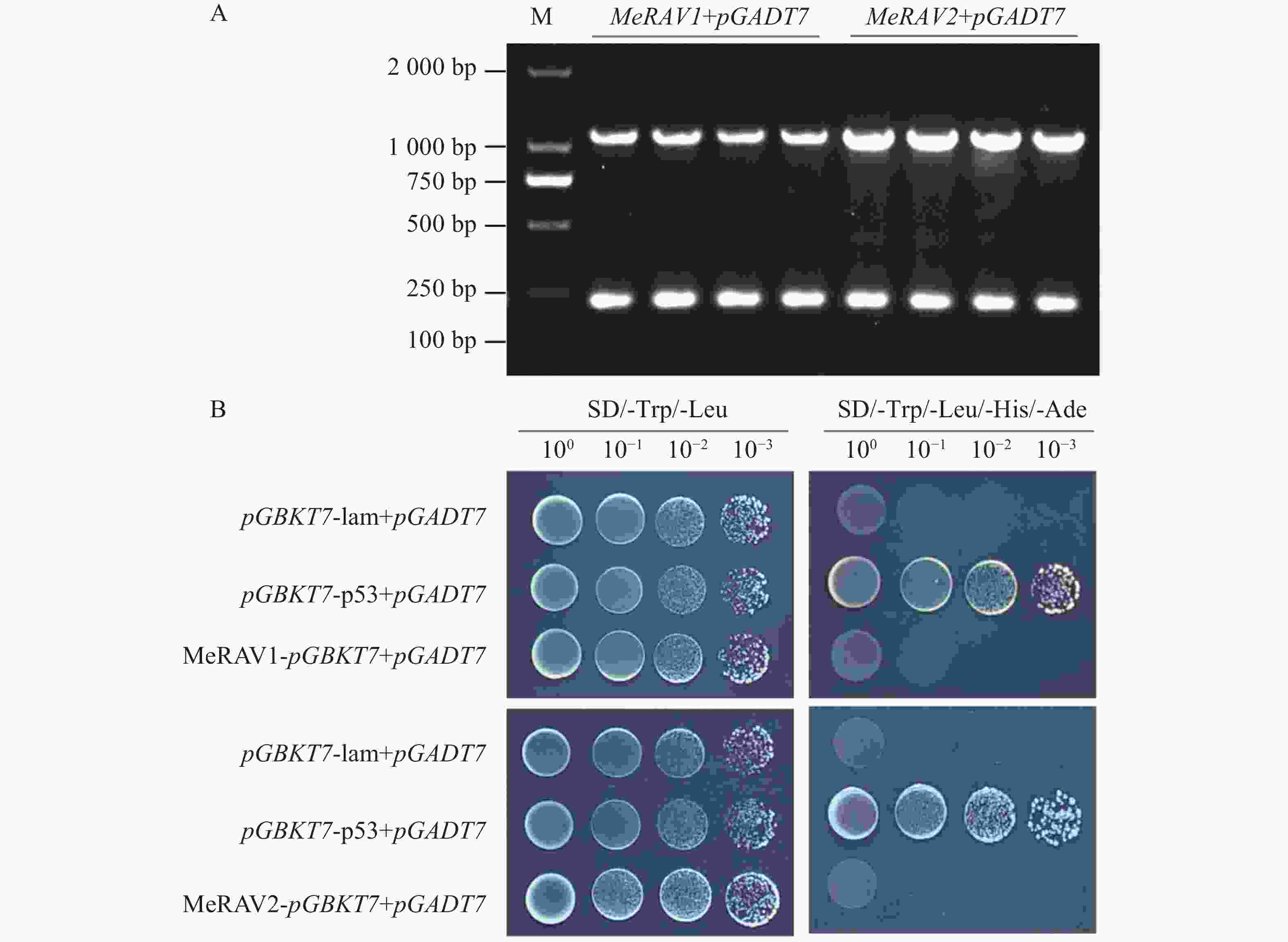
Figure 6. Self-activation assay of bait protein MeRAV1/2
-
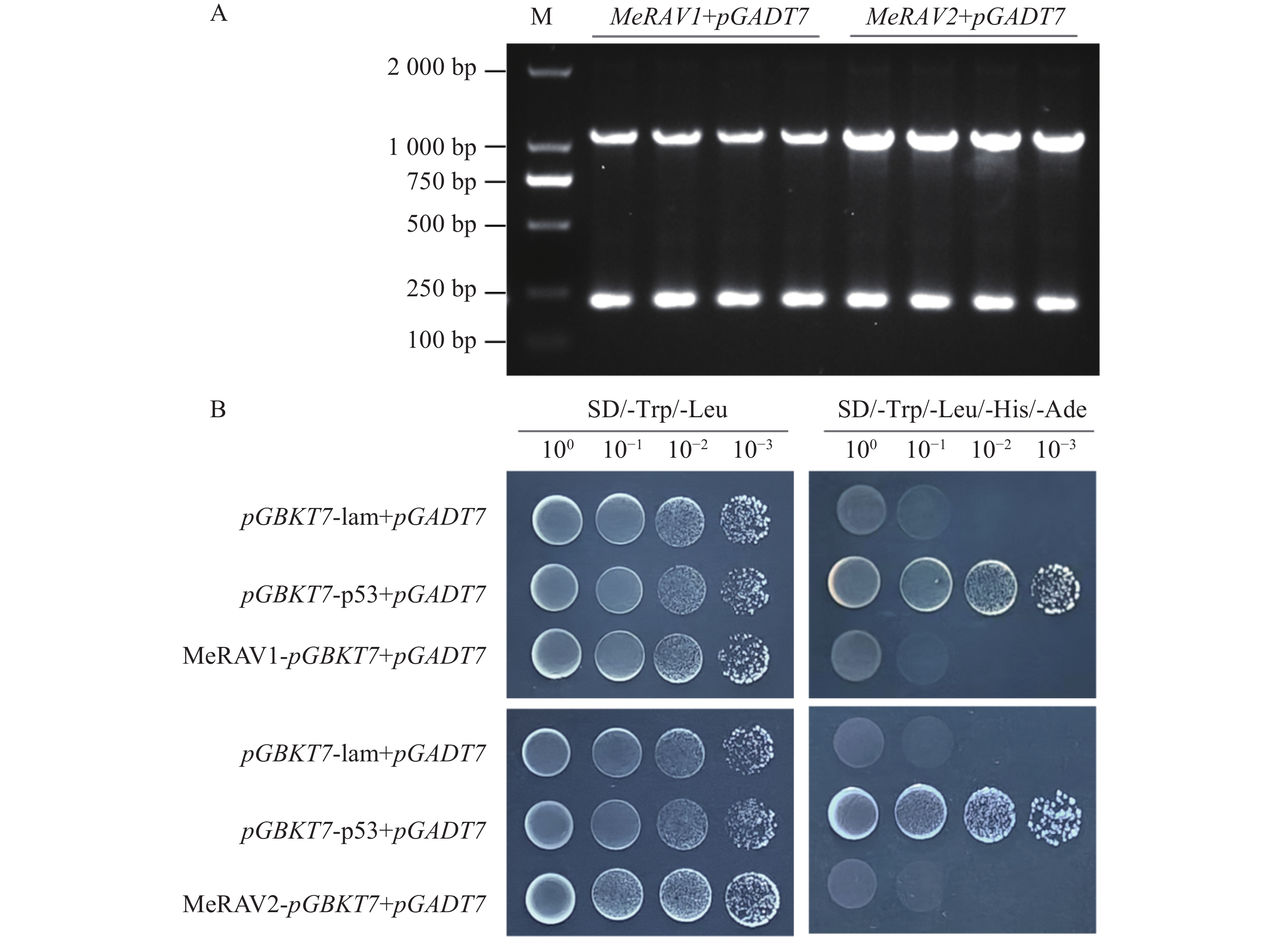
将MeKIN10全长和其3个结构域的猎物载体与诱饵载体pGBKT7-MeRAV1/2分别共转化至AH109酵母感受态细胞,在SD/-Trp/-Leu平板上,所有酵母菌株均可正常生长。随后,将阳性酵母克隆按不同梯度稀释并点滴在SD/-Trp/-Leu/-His/-Ade平板上,结果显示,除阳性对照和MeKIN10全长片段外,MeKIN10a可以和MeRAV1互作,MeKIN10a、MeKIN10b和MeKIN10c可以和MeRAV2互作(图7−A,B)。并进一步对其酵母菌株进行PCR检测,结果显示均为阳性(图8−A,B),且两条目的条带位置正确。以上结果表明MeRAV1仅与MeKIN10的第1个结构域STKc-AMPK-alpha存在互作关系,而MeRAV2与MeKIN10的3个结构域均具有互作关系。
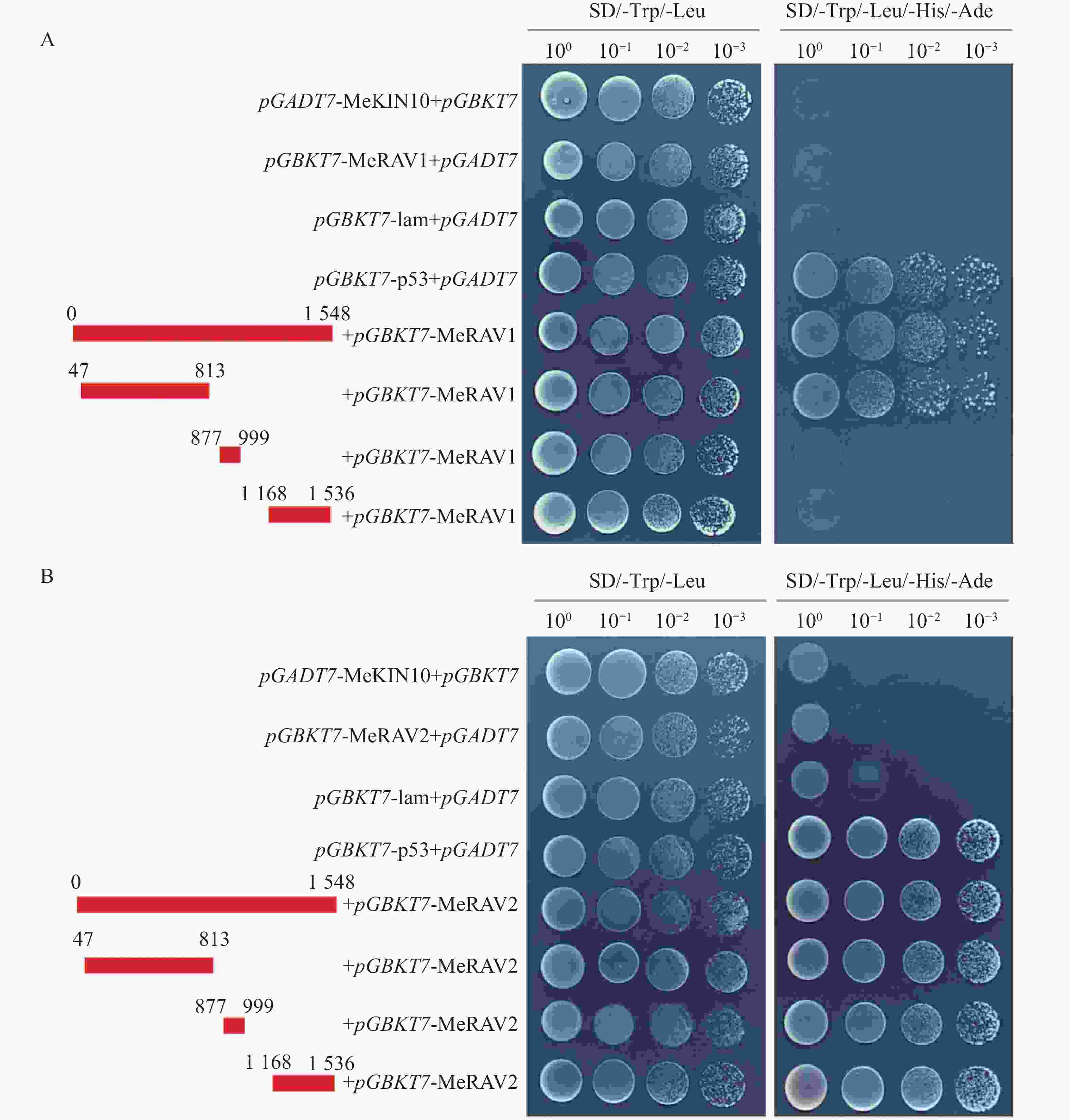
Figure 7. Verification of the interaction between MeRAV1/2 and MeKIN10
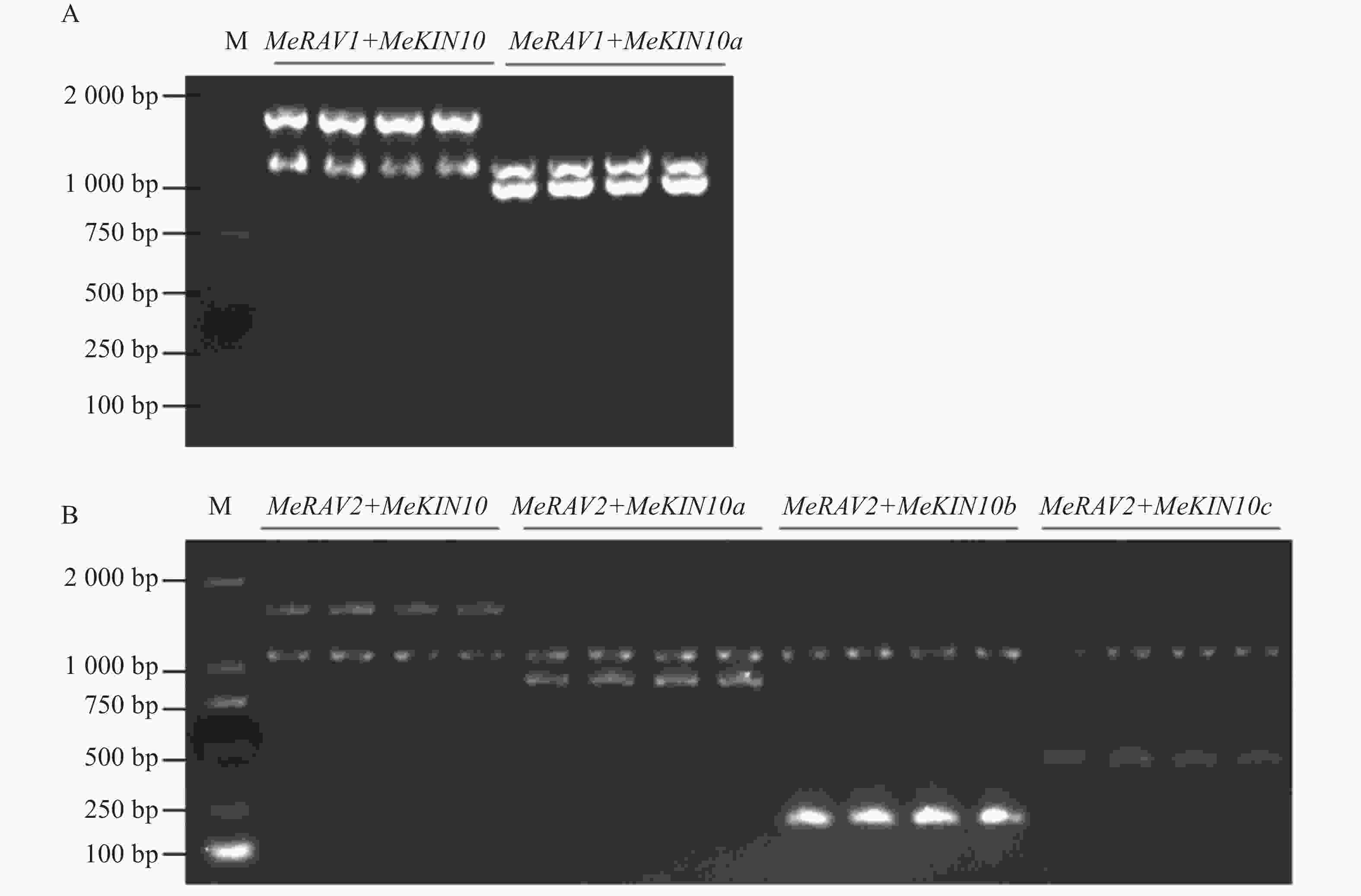
Figure 8. PCR verification of positive clones for MeRAV1/2 and MeKIN10
-
植物在面对逆境胁迫时需要不断调整自身的代谢从而适应多变的环境条件,SnRK1复合体是植物能量和环境胁迫信号的中心整合,它可以通过调节合成代谢和分解代谢的平衡来维持细胞能量稳态[14]。MeKIN10是SnRK1蛋白激酶行使磷酸化功能的1个重要α亚基,广泛参与了不同的逆境响应机制[15]。番茄中的SnRK1α1可以介导的RBOH1磷酸化调控ROS,增强番茄对低氮的耐受性[16]。木薯中的MeKIN10也有报道可以通过磷酸化MeHB16蛋白调控木质素合成,提高木薯的细菌性枯萎病抗性[17]。
本研究通过对MeKIN10与转录因子MeRAV1/2蛋白结构域分析表明,MeKIN10含有STKc-AMPK-alpha、UBA-SnRK1-plant及AMPKA-C 3个功能域,属于典型的SnRK1α蛋白。MeRAV1包含AP2和B3 DNA结合域,而MeRAV2额外具备KLF6-7-N-like superfamily结构域,表明其在基因调控网络中可能具有更复杂的功能。为了检测MeKIN10蛋白与MeRAV1/2蛋白相互作用的结构域,本研究构建了MeKIN10的全长序列与3个截断体MeKIN10a/b/c,以及MeRAV1/2的全长序列。可以根据这些截断体从而推测STKc-AMPK-alpha、UBA-SnRK1-plant及AMPKA-C 3个功能域中哪些结构域与MeRAV1/2的蛋白互作。
KIN10的激酶活性依赖于N端高度保守的激酶催化结构域,其展现出经典的二级折叠结构,含有一个激活环(T-loop),其关键氨基酸残基为参与磷酸集团转移反应的一个保守赖氨酸(K),突变形式的AtKIN10K48M不具有激酶活性[18 − 20]。在大麦中,SnRK1的激酶结构域能够磷酸化WRKY3,从而正向调节其对白粉病的抗性[21]。本研究的蛋白互作预测及酵母双杂交实验结果显示,MeKIN10的STKc-AMPK-alpha催化结构域与MeRAV1/2均互作,这说明其催化结构域可能是其介导磷酸化反应的关键。紧跟催化结构域之后,KIN10含有泛素相关(UBA)结构域,这是更广泛的AMPK相关激酶家族的共同特征[22]。在拟南芥中,已经有报道KIN10的UBA结构域与E3泛素连接酶复合物SKP1/ASK1相互作用,但其确切功能仍有待探索[23]。此外KIN10还具有C端调控结构域,参与调控SnRK其余亚基和上游磷酸酶的相互作用。在拟南芥中发现,ABI1和PP2CA能够与KIN10的AMPK-C结构域相互作用,导致其去磷酸化和失活[24]。本研究中发现,UBA-SnRK1-plant及AMPKA-C结构域只与MeRAV2互作。这表明MeKIN10通过其多个结构域与MeRAV2全面互作,可能在不同能量状态或环境条件下,赋予植物更灵活的氧化应激应答机制。然而,本研究采用的酵母双杂交系统存在异源表达的局限性,蛋白在酵母中的表达和折叠可能与植物细胞有所不同。因此,未来研究应结合植物本体的实验方法,如共免疫沉淀或双分子荧光互补,以进一步验证其互作区域。
综上所述,本研究鉴定了MeKIN10与MeRAV1/2蛋白互作的具体区域,进一步阐明了MeKIN10与MeRAV1/2的蛋白互作介导磷酸化的作用机制,并为深入探索植物应对能量需求变化及环境胁迫的分子机制提供了重要依据。
Identification of interaction domains between cassava MeKIN10 and MeRAV1/2 proteins
DOI: 10.15886/j.cnki.rdswxb.20250028
- Received Date: 2025-02-08
- Accepted Date: 2025-03-15
- Rev Recd Date: 2025-02-28
- Available Online: 2025-08-18
- Publish Date: 2025-09-25
-
Key words:
- cassava /
- KIN10 /
- RAV /
- protein domain /
- protein interaction
Abstract: Cassava (Manihot esculenta), a crucial staple and bioenergy crop, faces significant yield and quality losses due to oxidative stress induced by environmental factors such as drought and high temperature. Previous studies have demonstrated that MeKIN10, a protein kinase associated with energy sensing in cassava, enhances oxidative stress tolerance by mediating the phosphorylation of MeRAV1/2. While protein-protein interaction is essential for kinase-mediated phosphorylation, the specific interaction regions between MeKIN10 and MeRAV1/2 remain elusive. In this context the interaction regions of MeKIN10 and MeRAV1/2 were identified through protein domain analysis, AlphaFold3 prediction and yeast two-hybrid experiment. The results demonstrate that the STKc-AMPK-alpha kinase domain of MeKIN10 is the key domain in its interaction with MeRAV1/2, which further indicates that the protein interaction between MeKIN10 and MeRAV1/2 is the key in mediating the phosphorylation of MeRAV1/2.
| Citation: | HE Jiaoyan, CHEN Ao, YAN Yu. Identification of interaction domains between cassava MeKIN10 and MeRAV1/2 proteins[J]. Journal of Tropical Biology, 2025, 16(5): 663-672. doi: 10.15886/j.cnki.rdswxb.20250028

|
 DownLoad:
DownLoad: