
-
参薯(Dioscorea alata)为多年生缠绕藤本植物,其原产地分布于非洲、东南亚及南美洲热带地区[1]。中国规模化种植区集中在海南、云南、广西等地,具有重要的经济价值[2-3]。该物种的典型特征是形成肉质变态茎器官——块茎,通过储藏蛋白与淀粉的协同积累完成形态建成[4]。植物会受到多种胁迫,减产的主要因素是高温和干旱造成的[5]。同时植物为应对非生物胁迫进化形成了特定的代谢通路和调控系统。在维持正常生长发育节律、协调抗逆反应及病害防御机制,以及介导细胞信号传导网络对基因表达的调控等方面具有关键作用[6]。比如BBR-BPC成员在玉米(Zea mays)中的表达水平受温度、干旱等胁迫调控[7]。 通过基因表达调控提高参薯的抗旱、抗高温性等对参薯种植有着深远意义,在提高产量的同时扩大种植范围。Plant-specific Barley B Recombinant/Basic Penta Cysteine(BBR/BPC)家族成员是植物特异性GAGA基序结合因子(GAF),控制生长和繁殖的多个发育过程[8]。GAGA 基序结合因子(GAF)在许多多细胞生物体中具有不可或缺的正常生长和发育功能[9],在动物中,Trithorax like(Trl)和 Pipsqueak(Psq)蛋白家族是通过 DNA 环或与多梳抑制 DNA元件(polycomb repressive element, PRE)处的组蛋白修饰复合物的物理相互作用影响基因表达的 GAF[10-11]。尽管动物和植物蛋白构成不相关的蛋白质家族[12-13],但很早就提出它们在 PREs 中可能具有相同的分子功能[14]。BBR/BPC经常在拟南芥(Arabidopsis thaliana)基因的启动子中发现[15-16],在拟南芥基因组中 GA 重复序列的普遍性相吻合,BBR-BPC 调节广泛的同源基因以维持芽顶端分生组织的大小[17],根发育[15, 18],胚珠的发育转变[15, 19-22]。BPC基因功能缺陷引发拟南芥多维度发育表型:植株呈现显著矮化、叶片尺寸缩减伴顶端卷曲、胚珠发育异常等性状 [15]。近年研究发现,BBR-BPC转录因子家族在植物生长发育调控、非生物胁迫应答及激素信号转导过程中发挥重要作用。参薯作为重要的粮经兼用作物,其BBR-BPC基因家族的系统研究尚未见报道。
本研究用生物信息学分析参薯的基因结构,找到水稻(Oryza sativa)、拟南芥等模式植物的BBR-BPC 蛋白序列,构建系统进化树,同时对参薯 BBR-BPC的保守基序和表达模式进行分析。最后构建酵母双杂交相关载体,用酵母双杂交实验验证参薯 BBR-BPC 与参薯DELLA2的互作。
-
‘参薯70号’材料来自海南大学热带农林学院薯蓣实验室。PGBKT7-DELLA2相关载体由本实验室保存。本研究采用诺唯赞生物公司(南京)的FastPure系列试剂盒完成DNA凝胶回收、质粒提取等分子实验。DL2000 Plus DNA Marker、2×Rapid Taq Master Mix、2×Phanta Flash Master Mix也购买自诺唯赞生物公司(南京)。
-
多维度验证解析参薯BBR-BPC基因家族进化特征。基于拟南芥数据库(
https://www.arabidopsis.org/ )与NCBI(https://www.ncbi.nlm.nih.gov/ )数据库中获取拟南芥BBR-BPC蛋白序列、参薯 BBR-BPC 蛋白序列和现有参薯蛋白序列进行 BLAST 比对,去除重复序列。整合Phytozome v13(https://phytozome-next.jgi.doe.gov/ )平台中拟南芥、水稻、玉米、马铃薯(Solanum tuberosum)和新几内亚薯(Dioscorea rotundata)等物种的BBR-BPC蛋白序列构建多物种比对。为了进一步确定BPC基因成员,从Pfam数据库(http://pfam.xfam.org/ )中获得SRF-TF结构域(pfam accession: PF00319)的隐马尔可夫模型(HMMER),对参薯和其他5种上述物种的蛋白质进行搜索并进行BLAST 分析,核酸序列进行了多序列比对。得到的结果用MEGA v7.0基于邻接法(NJ)构建系统发育树(Bootstrap=1 000)。使用ExPASy-ProtParam(https://web.expasy.org/protparam/ )系统预测目标蛋白理化参数。 -
通过CD-Search(
https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi )分析6个DaBBR-BPC的共同保守结构。通过MEME Suite(https://meme-suite.org/meme/tools/meme )预测 参薯BBR-BPC保守基序位置。通过在线网站SOMPA(http://www.prabi.fr/ )和SWISS-MODEL(https://swissmodel.expasy.org )预测蛋白结构。 -
参薯BBR-BPC蛋白亚细胞定位分析使用在线软件WoLF PSORT(
https://wolfpsort.hgc.jp/ )。 -
Da4BBR-BPC2、Da5BBR-BPC和Da18BBR-BPC无内含子,利用 CTAB 法提取‘参薯 70号’DNA,以大薯 DNA 为模板扩增Da4BBR-BPC2、Da5BBR-BPC和Da18BBR-BPC。将 Da4BBR-BPC2、Da5BBR-BPC和Da18BBR-BPC构建到 PGADT7 载体上。分别命名为PGADT7-Da4BBR-BPC2、PGADT7-Da5BBR-BPC、PGADT7-Da18BBR-BPC。
将对照组PGBKT7-53+PGADT7-T和PGBKT7-lam+PGADT7-T和实验组1(PGBKT7-DaDELLA2//DaDELLA2-N/DaDELLA2-0.6K/DaDELLA2-1.2k/DaDELLA2突变体+PGADT7-Da4BBR-BPC2)和实验组2(PGBKT7-DaDELLA2/DaDELLA2突变体+PGADT7-Da5BBR-BPC、 PGBKT7-DaDELLA2/DaDELLA2突变体+PGADT7-Da18BBR-BPC)分别转入酵母感受态AH109,涂板在SD/-Leu/-Trp二缺板,30 ℃恒温培养2~3 d。
验证获得阳性菌液后,将OD600 调至0.2后稀释10、102、103倍,分别取实验组一以及相关对照组菌液3.5 μL点于SD/-Leu/-Trp、SD/-Leu/-Trp/-His/-Ade + 75 mmol·L−1 3-AT、SD/-Leu/-Trp/-His/-Ade、固体板上。取实验组二和对照组菌液3.5 μL点于SD/-Leu/-Trp、SD/-Leu/-Trp/-His/-Ade + 75 mmol·L−1 3-AT 固体培养基上,倒置于30 ℃恒温培养箱培养 3~5 d。
-
通过HMMsearch和BLAST分析以及Phytozome v13网站上下载拟南芥等模式植物BBR-BPC的表达序列,进行分析去除重复序列后在参薯基因组中鉴定了6个BBR-BPC家族成员,并分别命名为 Da4BBR-BPC1、Da4BBR-BPC2、Da5BBR-BPC、Da11BBR-BPC、Da14BBR-BPC和Da18BBR-BPC,它们的基因长度范围为 801~1 110 bp,氨基酸长度为267~370 aa(表1)。
基因编号
Gene ID基因名称
Gene name基因长度/bp
Gene length氨基酸数/aa
Number of amino acidsDioal.04G123900.1 Da4BBR-BPC1 1 110 370 Dioal.04G156300.1 Da4BBR-BPC2 822 274 Dioal.05G205200.1 Da5BBR-BPC 801 267 Dioal.11G001300.1 Da11BBR-BPC 954 318 Dioal.14G132600.1 Da14BBR-BPC 1 005 335 Dioal.18G083600.1 Da18BBR-BPC 816 272 Table 1. Structural information of the DaBBR-BPC gene
使用 CD-Search搜索DaBBR-BPC 基因的保守结构域,并对DaBBR-BPC基因家族进行进化关系的分析。结果显示,6个 DaBBR-BPC基因均含有共同的保守结构GAGA_bind(PF07859)。不同 DaBBR-BPC 转录因子的蛋白序列和理化性质存在较大差异(表2),结果显示,DaBBR-BPC基因家族分子质量为29 907.27~41 683.46 kDa; 该家族蛋白理论等电点介于9.39~9.84,具有典型碱性特征;亲水性分析显示平均系数均为负值,表明其亲水特性;不稳定指数(Ⅱ)均高于40,提示为不稳定蛋白;脂肪系数在57.43~68.24。
基因名称
Gene name分子质量/kDa
Molecular
Weight等电点
Theoretical pI亲水性平均系数
Grand average of
hydropathicity不稳定指数
Instability
index脂肪系数
Aliphatic indexDa4BBR-BPC1 41683.46 9.49 −0.798 52.92 63.62 Da4BBR-BPC2 30292.57 9.82 −0.720 43.17 60.15 Da5BBR-BPC 29907.27 9.77 −0.723 50.43 61.76 Da11BBR-BPC 35429.32 9.39 −0.695 53.21 68.24 Da14BBR-BPC 37730.96 9.62 −0.851 41.35 58.93 Da18BBR-BPC 30231.76 9.84 −0.669 49.61 57.43 Table 2. Analysis of physicochemical properties of DaBBR-BPC gene
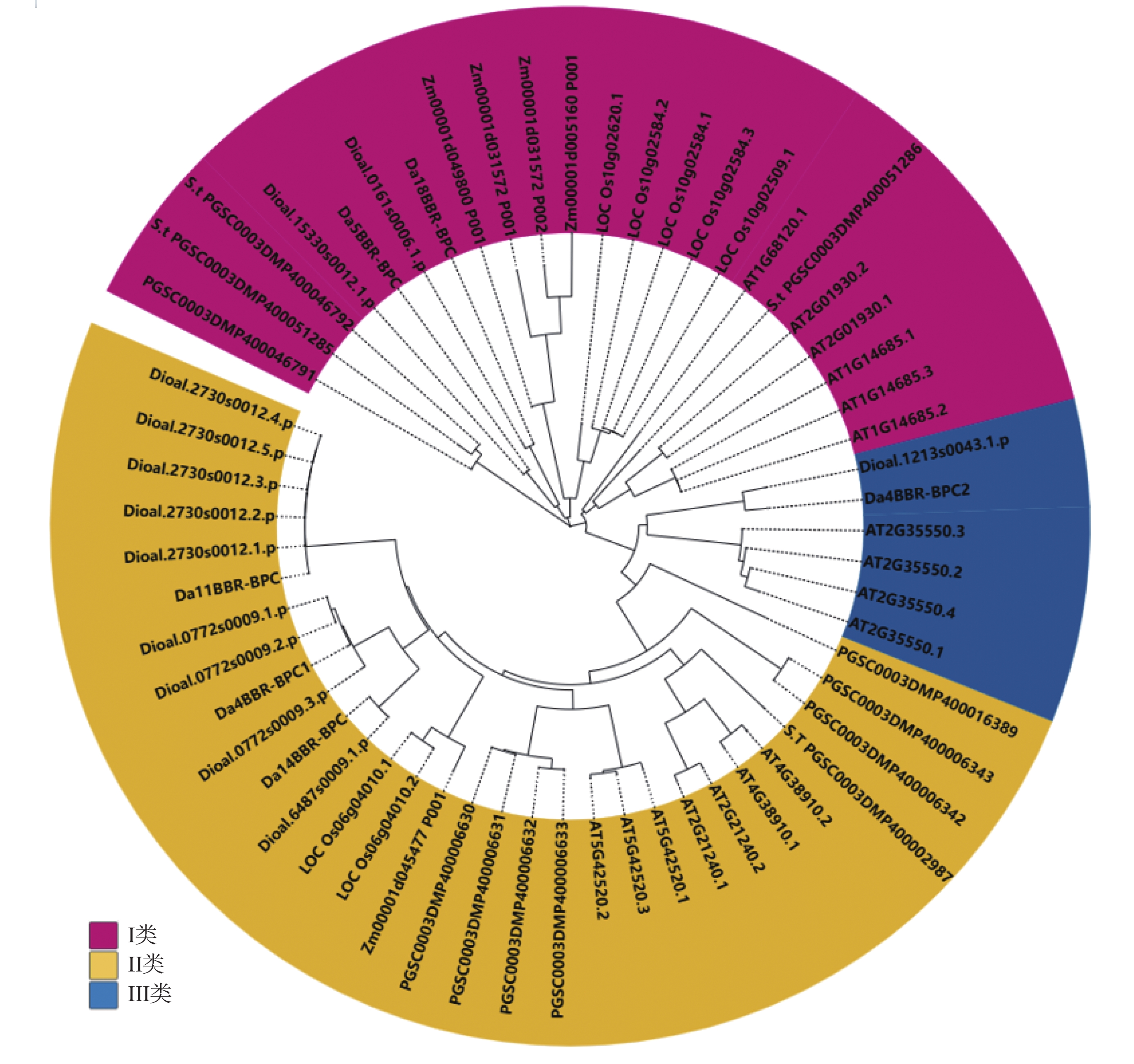
根据Phytozome v13网站下载的参薯、拟南芥和马铃薯等植物的蛋白序列,用 MEGA7进行多序列比对,并构建相关系统进化树。从图1可知,在拟南芥中,有7个广泛表达的成员(BPC1-7),拟南芥基因组序列编码的 7 个 BPC 分为3类,即Ⅰ类 (BPC1~3)、Ⅱ 类 (BPC4~6)和Ⅲ类 (BPC7)。除了被认为是假基因的BPC5外,它们都是普遍表达的转录激活因子和抑制因子[15, 19]。根据拟南芥BPC基因的家族分类,把参薯BBR-BPC亚家族也分为3类,Da5BBR-BPC和Da18BBR-BPC为Ⅰ类,Da4BBR-BPC1、Da11BBR-BPC和Da14BBR-BPC为Ⅱ类,Da4BBR-BPC2为Ⅲ类。
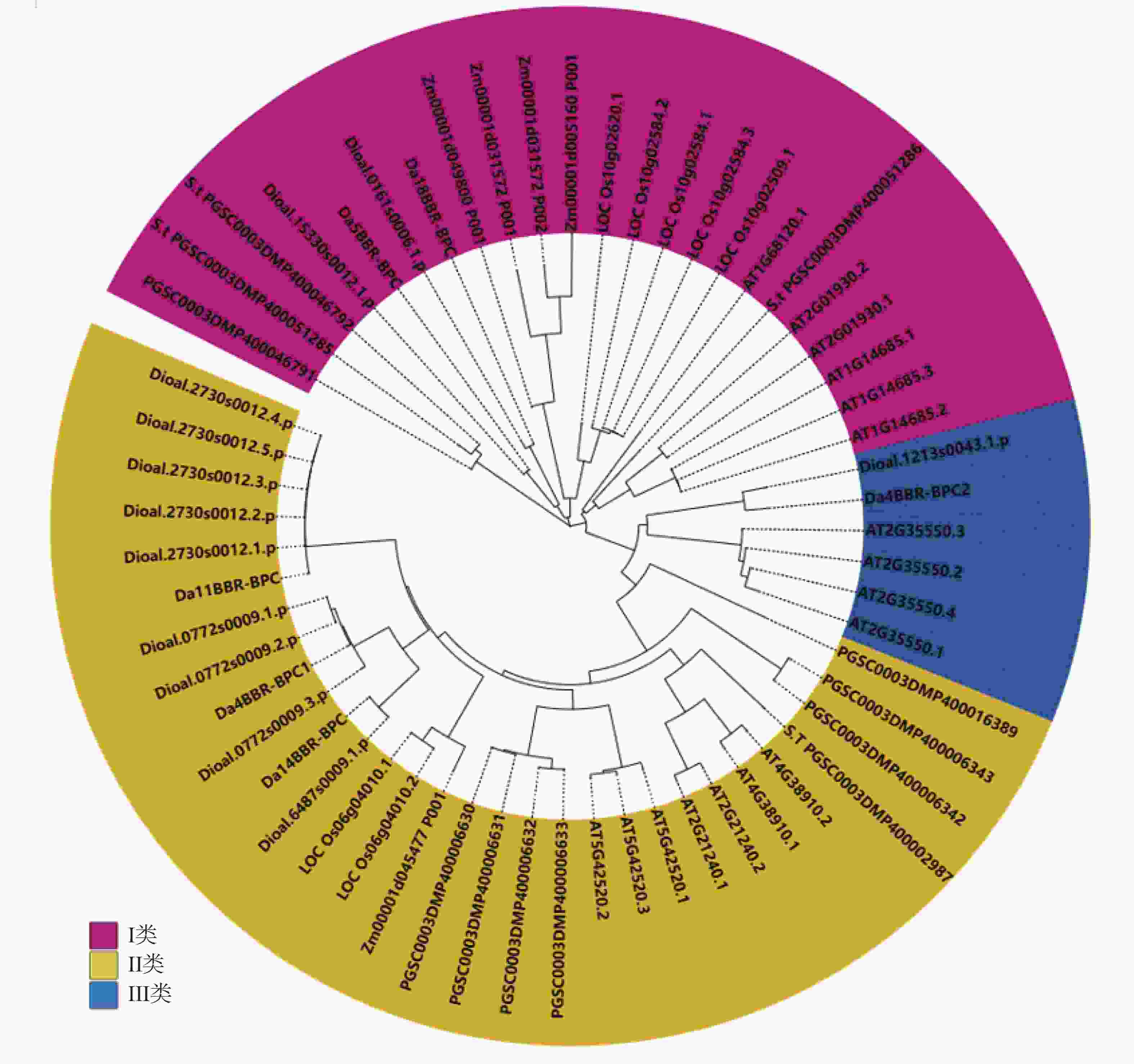
Figure 1. Phylogenetic tree analysis of BBR-BPC gene family
-
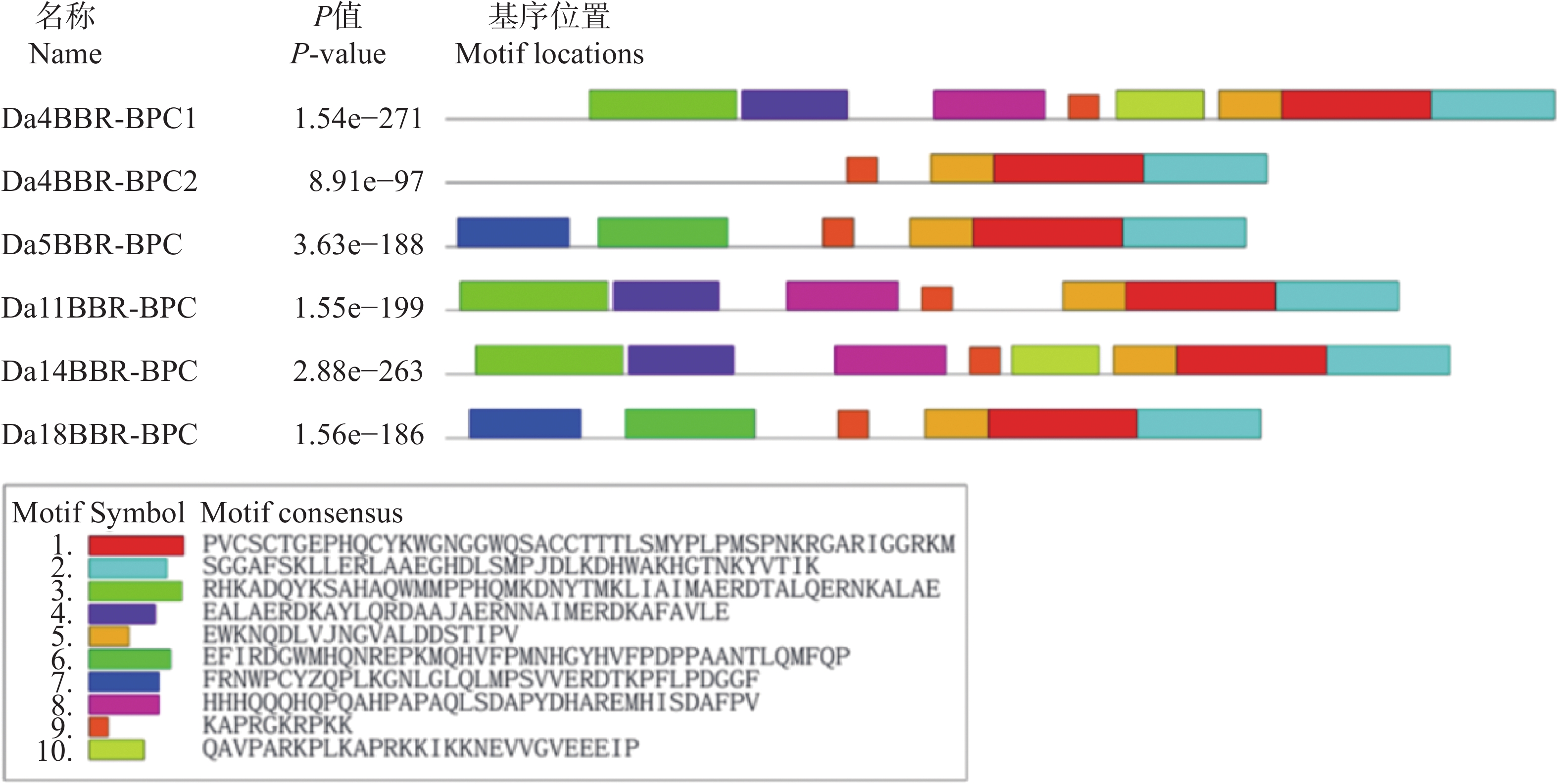
基于MEME Suite对参薯BBR-BPC进行保守基序解析(图2)。结果显示,核心基序(motif1/2/5/9)在6个家族成员中呈现完全保守性。
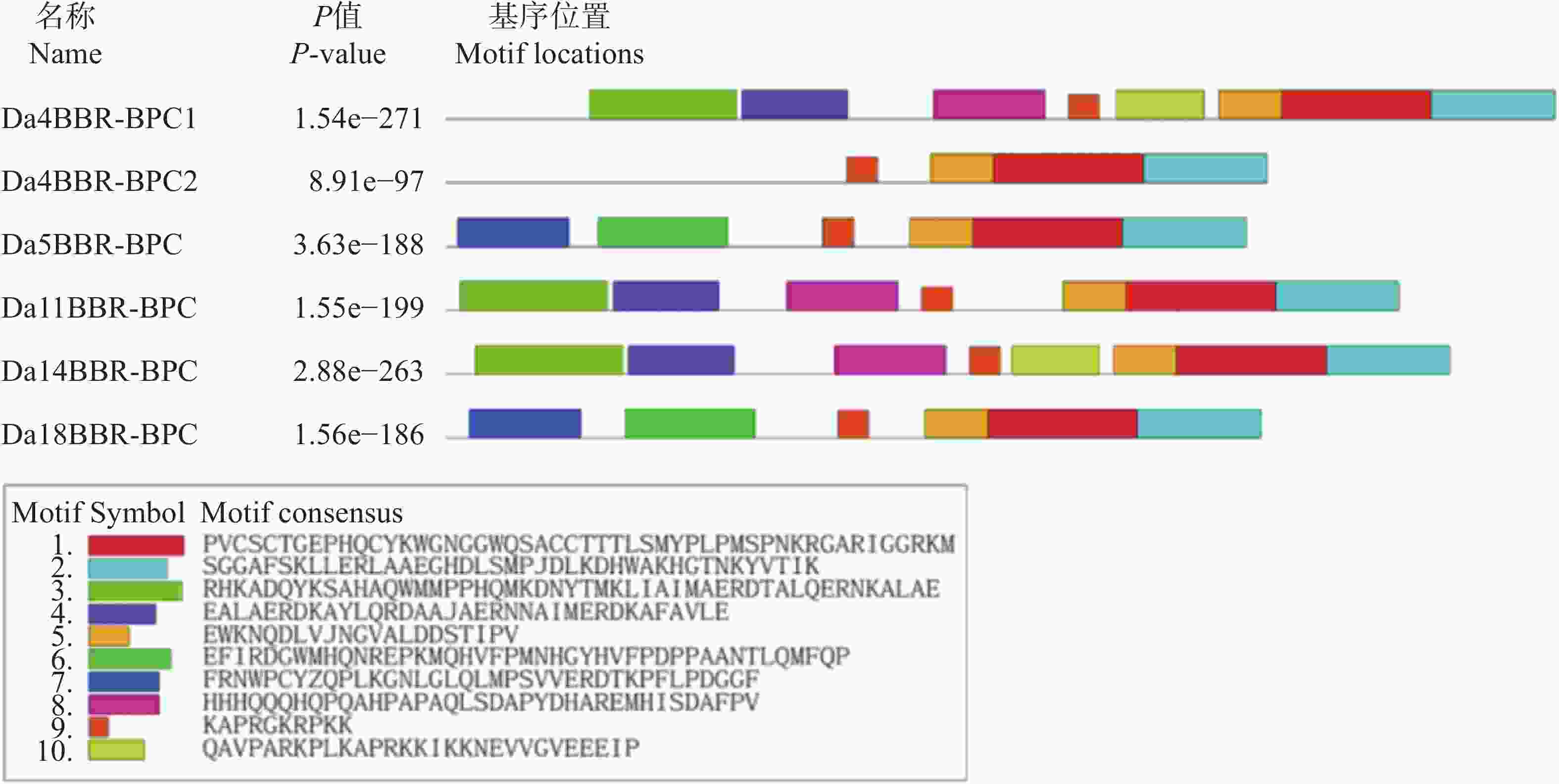
Figure 2. Distribution of conserved motifs on Dioscorea alata BBR-BPC protei
-
结果显示(表3),Da4BBR-BPC1、Da4BBR-BPC2、Da5BBR-BPC、Da11BBR-BPC、Da14BBR-BPC、Da18BBR-BPC在细胞核中都有分布;Da11BBR-BPC、Da14BBR-BPC在细胞质中有分布,Da4BBR-BPC1、Da11BBR-BPC和Da18BBR-BPC在叶绿体中有分布;Da4BBR-BPC1、Da11BBR-BPC、Da14BBR-BPC和Da18BBR-BPC在线粒体中有分布;Da4BBR-BPC2分布在细胞核中,在细胞质、叶绿体和线粒体中均没有分布;Da11BBR-BPC在细胞核、细胞质、叶绿体和线粒体中均有分布。
基因名称
Gene name细胞核
Nucleus细胞质
Cytoplasm叶绿体
Chloroplast线粒体
MitochondriaDa4BBR-BPC1 8.0 0 4 1 Da4BBR-BPC2 14.0 0 0 0 Da5BBR-BPC 14.0 0 0 0 Da11BBR-BPC 7.5 4 1 1 Da14BBR-BPC 8.0 2 0 3 Da18BBR-BPC 11.0 0 1 1 Table 3. Subcellular localization prediction of DaBBR-BPC
-
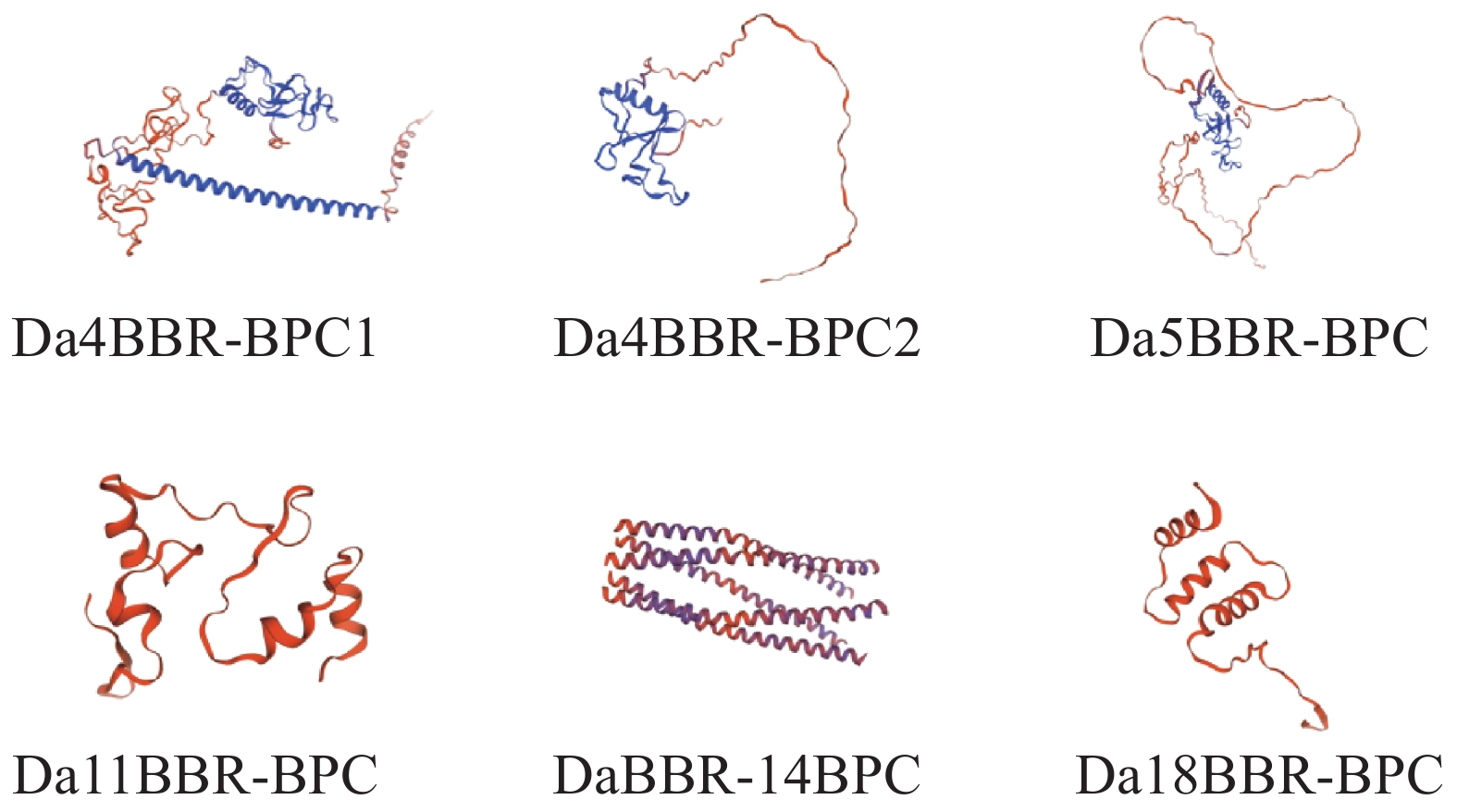
通过生物信息学分析平台SOMPA和SWISS-MODEL对DaBBR-BPC蛋白进行结构预测。二级结构解析表明(表4),无规则卷曲(30.45%~68.54%)和α−螺旋(21.32%~64.18%)构成主要结构元件,两者合计占比超过88%,而延伸链(2.69%~7.72%)和β−转角(2.69%~4.04%)比例较低,未见β−折叠结构。三级结构建模显示(图3),6个家族成员具有高度保守的空间构象。
名称
Nameα−螺旋/%
Alpha helixβ−折叠
Beta sheetβ−转角/%
Beta turn无规则卷曲/%
Random coil延伸链/%
Extended strandDa4BBR-BPC1 57.03 0 3.24 34.86 4.86 Da4BBR-BPC2 29.2 0 3.65 60.22 6.93 Da5BBR-BPC 21.72 0 3.00 68.54 6.74 Da11BBR-BPC 53.14 0 3.46 38.68 4.72 Da14BBR-BPC 64.18 0 2.69 30.45 2.69 Da18BBR-BPC 21.32 0 4.04 66.91 7.72 Table 4. Secondary structure analysis of DaBBR-BPC protein
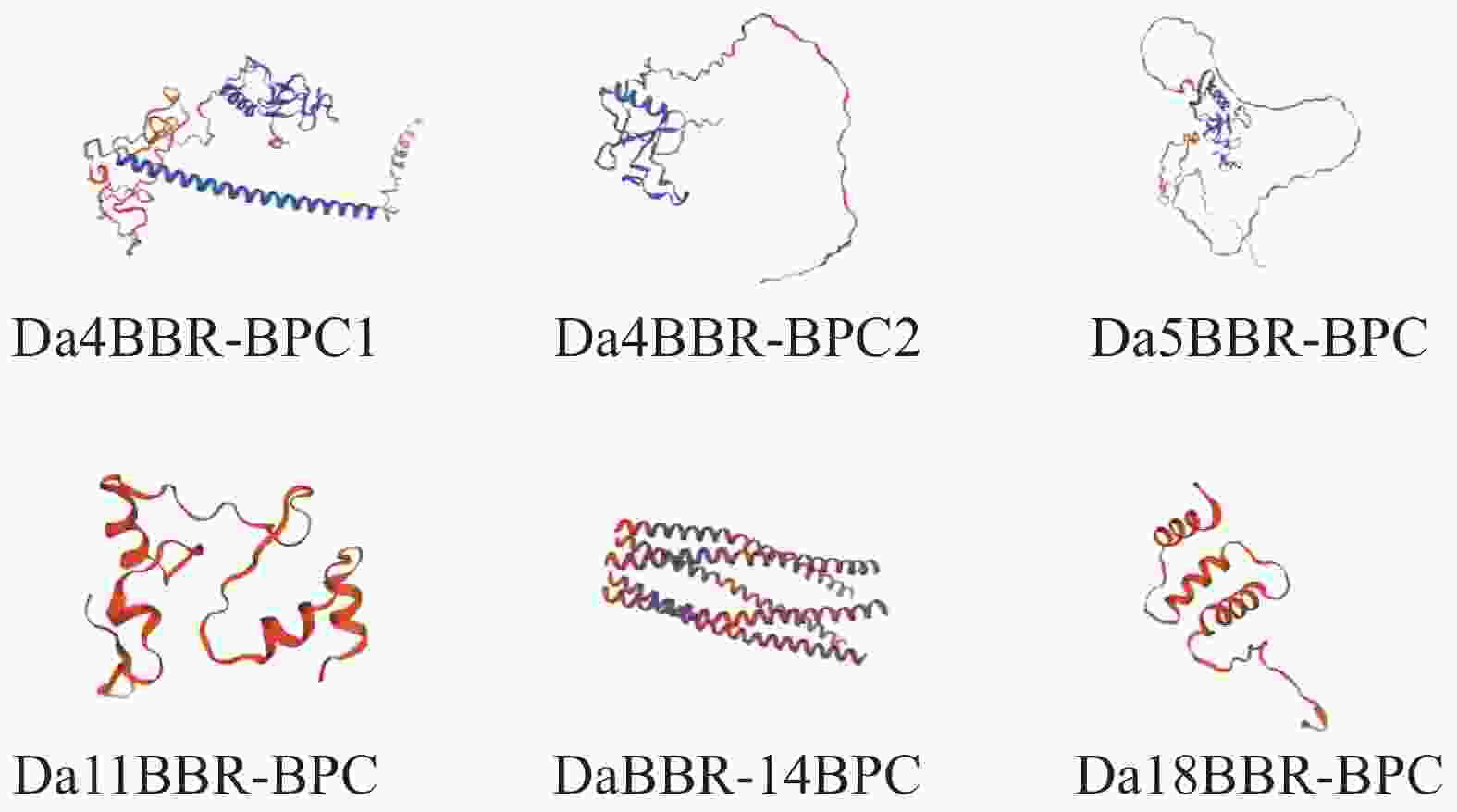
Figure 3. Prediction of tertiary structure of DaBBR-BPC protein
-
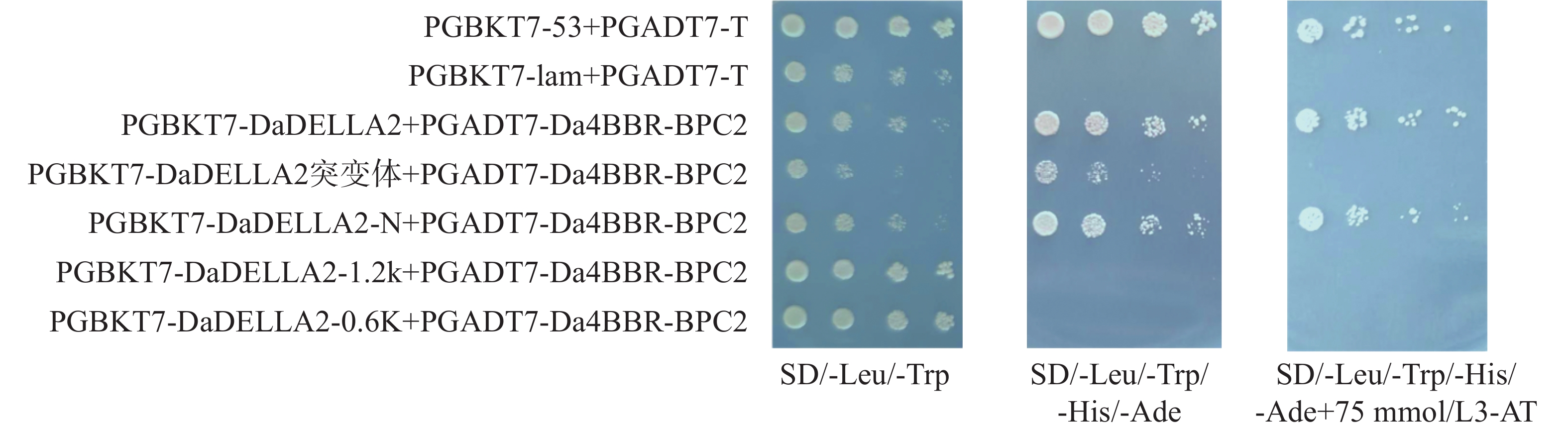
基于实验室前期建立的75 mmol·L−1 3-AT浓度下抑制参薯 DELLA2 蛋白自激活体系,将对照组和实验组分别转入酵母感受态AH109,验证阳性菌液后将OD600 调至0.2,采用梯度稀释点板法,稀释后点板。结果显示(图4),PGBKT7-DaDELLA2/DaDELLA2-N+PGADT7-Da4BBR-BPC2转化后的酵母菌能在 75 mmol·L−1 3-AT的四缺板上长出。结果表明,DaDELLA2 与 Da4BBR-BPC2存在互作,且互作位置发生在DaDELLA2蛋白的 N端。同理将对照组和实验组二转入酵母感受态AH109;验证阳性菌液后调节OD600 至0.2,并分别稀释后点于相应的固体板上。酵母菌生长结果显示(图5),PGADT7-Da5BBR-BPC、PGADT7-Da18BBR-BPC与DaDELLA2也存在互作。
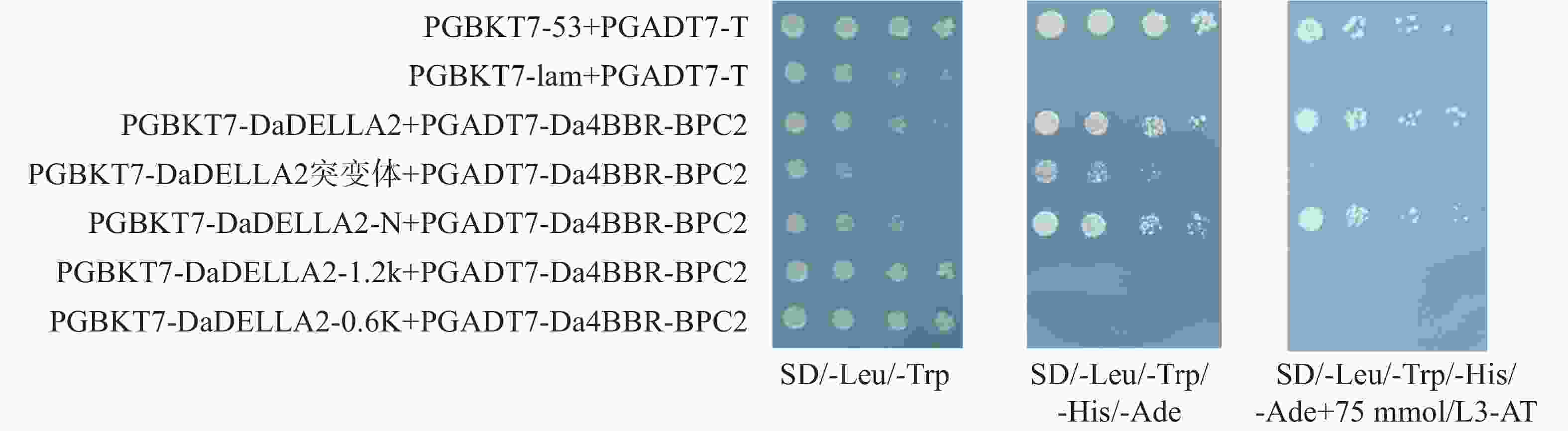
Figure 4. Detection of DaDELLA2 and Da4BBR-BPC2 protein interaction
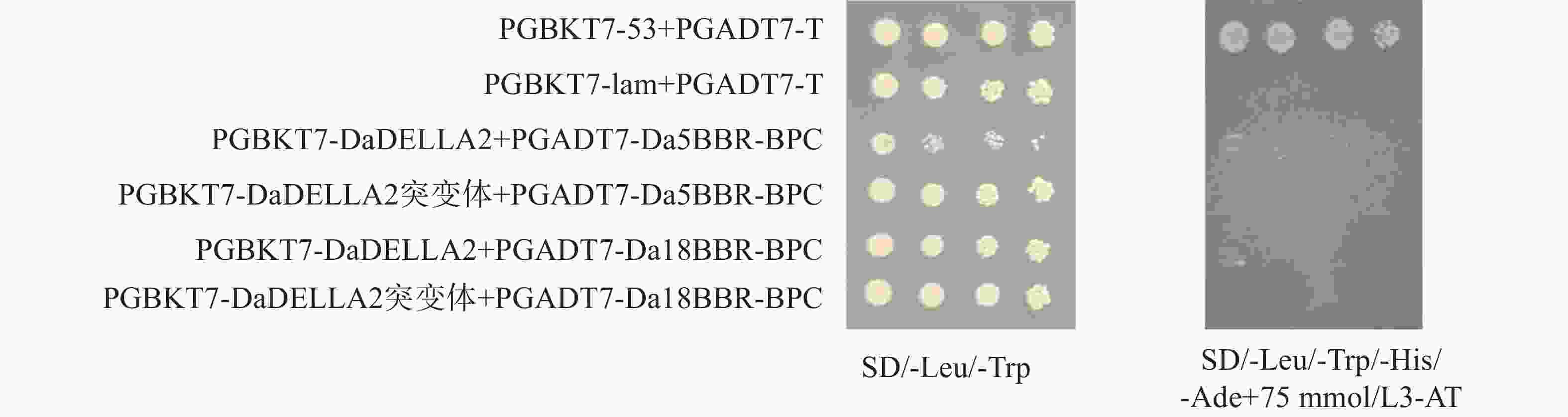
Figure 5. Detection of DaDELLA2 and Da5BBR-BPC、Da18BBR-BPC protein interaction
-
碱性五半胱氨酸(BPC)转录因子的植物特异性家族参与植物发育各个方面,包括胚胎发育、根生长和开花[15, 20]。大麦(Hordeum vulgare)B重组/碱性五半胱氨酸 (BBR-BPC)蛋白家族是与 GAGA 基序结合的最小转录因子之一[23-24]。黑腹果蝇(Drosophila melanogaster)GAGA 因子(dGAF)与这2种类型的 Chromatin-associated proteins(CAP) 相互作用以调节同源框基因[25]。CsGAox 基因启动子的候选转录因子 BBR/BPC和 GRAS(赤霉酸不敏感区)[26]。研究发现,BBR-BPC的基因在植物中基因组数量较少,在单子叶参薯、水稻等作物中存在6~8个BBR-BPC基因,有的作物甚至更少[27]。
BBR-BPC可能是调控草地早熟禾(Poa pratensis)根茎发育的关键转录因子[27]。BBR-BPC是NAC(NAM、ATAF1/2和CUC2)的潜在TFs,在干旱胁迫条件下,参与百香果(Passiflora edulis)的调控[28]。BBR/BPC TF 家族的标志是存在1个具有 5 个半胱氨酸残基的保守 C 端区域[27]。BBR-BPC是预测的RNAi(DCL、AGO和RDR)相关基因的关键转录调节因子, 小麦(Triticum aestivum)RNAi 基因对干旱和高温胁迫有显著反应[29]。BPCs可以通过结合GA基序来调节靶基因的表达,表明BPC s的表达模式可能影响其时空调控[22]。在椰子(Cocos nucifera)中,同一类CnBPC s的表达模式也非常相似,从而确保了它们的功能角色重叠[22],研究表明,BPC基因具有功能冗余性[15]。本研究分析参薯的BBR-BPC家族,结果发现6个DaBBR-BPC基因,它们均含有共同的保守结构域GAGA_bind(PF07859),且不同 DaBBR-BPC 转录因子的蛋白序列和理化性质存在差异,但均属于碱性、亲水性,属不稳定蛋白。Da4BBR-BPC2只分布在细胞核中,Da11BBR-BPC在细胞核、细胞质、叶绿体和线粒体中均有分布。
参薯营养价值丰富、色泽鲜艳、口感俱佳,是人们喜爱的一种食品[30]。赤霉素(Gibberellin,GA)在块茎发育过程中的作用被广泛研究,赤霉素的应用促进了匍匐茎的伸长并抑制了块茎的形成[31]。同时,植物赤霉素信号传导途径中的负调控蛋白为DELLA蛋白[32]。在赤霉素信号转导通路中,DELLA蛋白作为植物特异性调控因子展现出双重核心功能:其一通过动态负反馈调节机制,直接作用于赤霉素生物合成关键酶编码基因及其受体基因的转录活性,从而精准维持胞内赤霉素稳态;其二通过激活E2/E3泛素连接酶复合体及相关转录调控因子的生物合成,有效增强泛素–蛋白酶体降解级联反应中的负调控元件活性[33]。通过酵母实验发现,DaDELLA2 与 Da4BBR-BPC2、Da5BBR-BPC和Da18BBR-BPC存在互作关系,且在与Da4BBR-BPC2酵母实验结果中发现,Da4BBR-BPC2发生在DaDELLA的 N端,表明参薯中的BBR-BPC基因对DELLA2起到调控作用。BPCs与其他转录因子和染色质重塑蛋白的相互作用进一步复杂化了其调控机制[34-35]。所以,BBR-BPC 家族和赤霉素在参薯生长调控和结薯过程中有着重要的作用,研究它们之间的关系有利于更好地调控参薯的生殖生长,从而产生更高的经济效益。
Identification and characteristic analysis of BBR-BPC gene family in greater yam (Dioscorea alata L.)
DOI: 10.15886/j.cnki.rdswxb.20250026
- Received Date: 2025-02-04
- Accepted Date: 2025-02-28
- Rev Recd Date: 2025-02-22
- Available Online: 2026-05-20
- Publish Date: 2026-05-25
-
Key words:
- DaBBR-BPC gene /
- DaDELLA2 /
- Gibberellin /
- Yeast two-hybrid /
- Bioinformatics analysis
Abstract: In order to understand the regulatory role of the BBR-BPC gene family in the growth and development of Dioscorea alata (greater yam), we analyzed the gene structure, phylogenetic evolution, conserved motifs, and physicochemical properties of the BBR-BPC gene family in greater yam. We also predicted the secondary and tertiary structures of BBR-BPC proteins and verified the interaction between the BBR-BPC gene and the DELLA gene. Through HMMsearch and BLAST analyses, we identified six members of the BBR-BPC family in the greater yam genome. These gene proteins exhibited significant differences and alternative splicing events, with a total of 10 splice variants detected. Subcellular localization analysis revealed that all members of the BBR-BPC family in greater yam are located in the nucleus.The yeast two-hybrid interaction results between Da4BBR-BPC2 and DaDELLA2 indicate that there is an interaction between Da4BBR-BPC2 and DaDELLA2, occurring at the N-terminus of the DaDELLA2 gene.
| Citation: | Zhao Aote, Lin Huiting, Zhang Yuting, Huang Xiaolong, Wu Wenqiang, Xia Wei, Huang Dongyi. Identification and characteristic analysis of BBR-BPC gene family in greater yam (Dioscorea alata L.)[J]. Journal of Tropical Biology, 2026, 17(3): 390-398. doi: 10.15886/j.cnki.rdswxb.20250026

|
 DownLoad:
DownLoad: