
-
实时荧光定量PCR(reverse transcription quantitative real-time PCR, qRT-PCR)是基因表达研究中最准确、最可靠的技术之一[1-2]。该方法通过荧光信号测定PCR 循环扩增产物总量,并利用内参基因作为参考对特定cDNA序列进行定量分析[3-4]。qRT-PCR常用于检测基因在不同处理、不同生长发育时期的表达量变化,而内参基因的稳定性决定了检测基因表达量变化的准确性。因此,使用稳定表达的内参基因对目的基因表达水平的标准化至关重要[5]。好的内参基因参与细胞的基础代谢功能,并在整个过程中保持稳定表达,不受外部条件改变的影响[6-7]。然而,基因在生物体中不同组织和不同环境下行使功能的差异会导致没有任何一个内参基因能够在所有条件下都恒定表达[8]。因此,应根据具体的实验条件筛选出表达量相对稳定的内参基因来对靶标基因的表达进行分析,从而获得可靠的试验结果。
木薯(Manihot esculenta),是大戟科(Euphorbiaceae)木薯属植物,与马铃薯(Solanum tuberosum )、红薯(Ipomoea batatas )并列为世界重要的薯类作物,被誉为“地下粮仓”和“淀粉之王”[9-11]。木薯具有多种优良特性,如耐瘠、耐旱、适应性强、管理粗放、容易栽培和淀粉含量高等特点[12];块根淀粉含量丰富且种植易成活的优势,使其成为了热带地区低收入国家的主要人类粮食[13]。木薯还能作为乙醇的主要生产原料,也可以用以制作沼气、纺织、医疗药品等[13-14]。但病害是制约木薯产业健康、持续发展的重要因素之一。世界范围内,危害木薯生产的病害高达30多种,其中木薯细菌性枯萎病(cassava bacterial blight,CBB)的危害尤其严重[15]。CBB是由菜豆黄单胞菌木薯萎蔫致病变种(Xanthomonas phaseoli pv. manihotis,Xpm)引起的检疫性病害,导致木薯严重减产甚至绝收[16-17]。尽管对CBB的研究已取得一定进展,但相关研究大多停留在生理水平,其在分子层面上的抗病机制仍有待进一步阐明。目前,尚未有木薯在Xpm侵染条件下稳定表达内参基因的研究报道,因此,筛选在Xpm侵染木薯过程中稳定表达的内参基因是研究木薯响应细菌性枯萎病相关功能基因定量表达准确性的关键因素,可为木薯抗病分子育种基因功能研究提供衡量基因表达水平的标尺。
-
木薯抗病品种‘JG1301’、感病品种‘SC9’和菜豆黄单胞菌Xpm均由海南大学热带农林学院热带作物与微生物互作团队提供。
-
随机选择生长良好、高度和长势一致的3株生长45 d的木薯‘JG1301’和‘SC9’,采用叶片注射法接种病原菌Xpm,每个品种接种3株,每株接种3个掌叶,每片掌叶接种2个复叶;接种后分别于0、6、12、72和120 h 采取叶片,每个时间点从3株木薯各采1复叶(3次重复),采集后放入离心管中,液氮速冻后,−80 ℃保存备用。
-
使用天根RNAprep Pure多糖多酚植物总RNA提取试剂盒(DP432)按照说明书方法进行木薯叶片总RNA的提取。用微型分光光度计(Nano−300 Micro-Spectrophotometer)检测RNA浓度和质量。通过1.0%琼脂凝胶电泳检测RNA完整度。采用天根FastKing一步法除基因组cDNA第一链合成预混试剂(KR118),按照说明书方法进行反转录,使用1 μg总RNA建立20 μL cDNA逆转录反应体系。反转录后的cDNA于−80 ℃冰箱保存备用。
-
根据文献[15, 18-22]查找木薯研究中qRT-PCR实验常用的内参基因,并最终筛选出实验所需要的 5个内参基因,分别为TUB、Nascent、GTPb、ACT和EF1a;通过在Phytozome数据库中查询基因序列,确定各候选基因的CDS序列,根据基因的CDS序列使用在线工具Primer3 web(
https://primer3.ut.ee/ )进行引物设计;通过Phytozome Blast鉴定引物特异性后,送北京擎科生物科技股份有限公司进行引物合成,引物序列见表1。基因符号
Gene引物序列(5′−3′)
Primer sequences (5′−3′)基因登录号
Accession No.扩增长度/bp
Length引物扩增效率/%
Efficiency决定系数R2
Correlation R2参考文献
ReferencesTUB F: gtggaggaactggttctgga
R: tgcactcatctgcattctccManes.08G061700 180 105.5 0.997 [15] Nascent F: ttttgaagctgctgcagagg
R: tgaagcggatctgaacaaccManes.09G005100 192 100.7 0.997 [18] GTPb F: ggtgggtaacaaggcagaca
R: tgagtcggtatctgcaagccManes.09G086600 169 102.4 0.993 [19] ACT F: ttcgtgtcaaggtgtcgtga
R: gccctctcatttgctgcaatManes.13G086400 149 106.7 0.999 [20] EF1α F: tgaaccaccctggtcagattggaa
R: aacttgggctccttctcaagctctManes.15G054800 139 109.4 0.998 [21 - 22] Table 1. Reference genes and their corresponding primers
-
对每一对内参基因引物制作对应的标准曲线,计算相应引物的扩增效率。以ddH2O作为阴性对照,用于检测实验过程中的试剂或者人为污染。所有样品重复3次。采用耶拿荧光定量PCR qTOWER384G(Analytik Jena)进行qRT-PCR实验,根据测定的Ct值和对应的不同稀释梯度模板浓度经线性回归(其斜率值即为 slope),计算得各候选基因的引物扩增效率(E=10−1/slope)及决定系数R2。
-
采用耶拿荧光定量PCR qTOWER384G(Analytik Jena)进行qRT-PCR实验,每个反应均包含正向和反向引物各2 μL(10 μmol·L−1),不同稀释倍数的cDNA模板5 μL(20 ng·μL−1),10 μL 2× SYBR Greenq PCR Mix(2×SYBR Green qPCR Mix,思科捷),加 ddH2O至20 μL反应体系。 每个反应体系进行3次重复。qRT-PCR 反应条件:94 ℃预变性 3 min,随后进行 94 ℃变性10 s,60 ℃延伸34 s,循环40次的二步法程序。在每个实验结束时,使用默认参数在60~94 °C以0.3 ℃的增量持续60 s的周期进行熔解曲线分析。
-
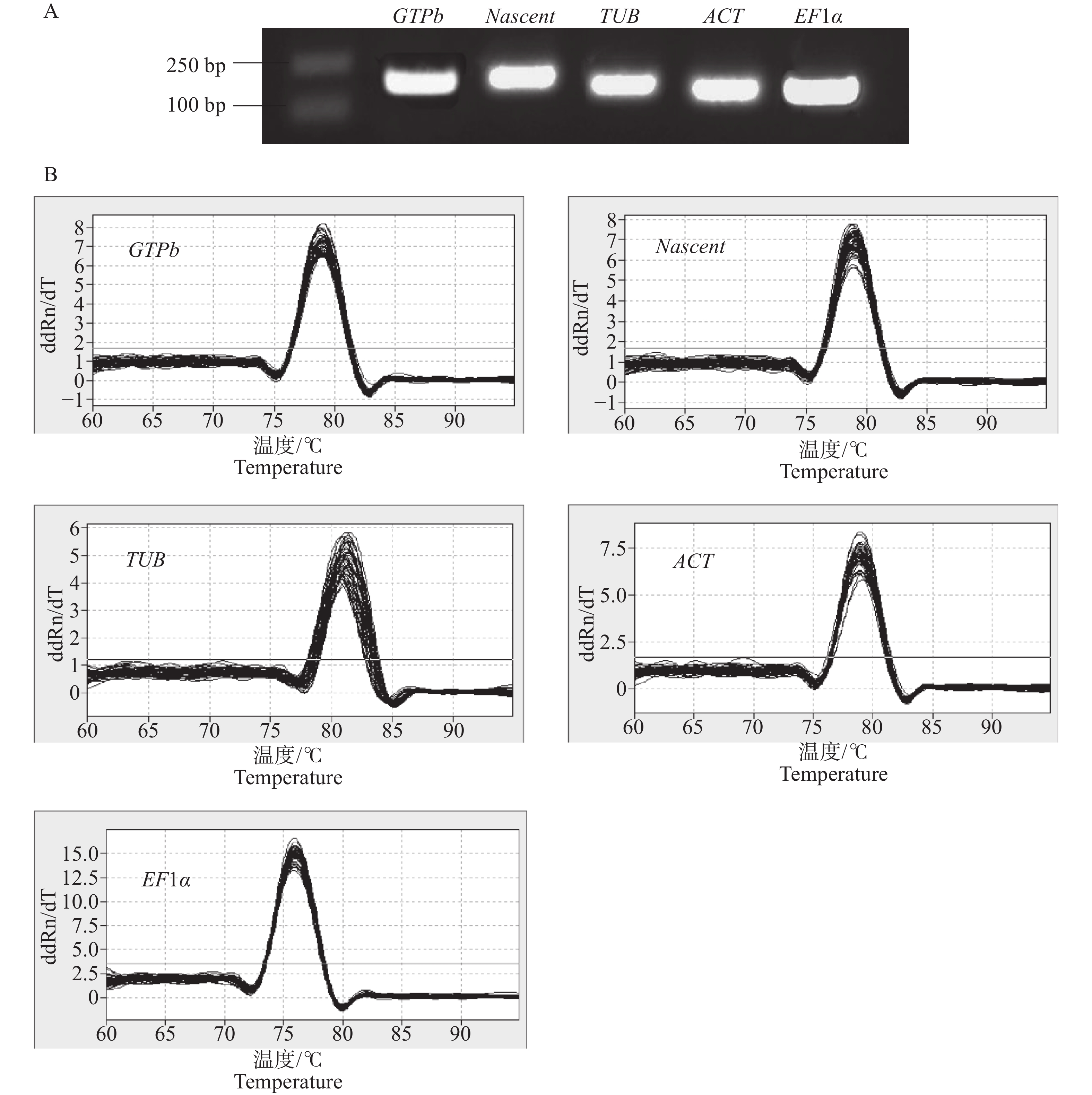
以Xpm侵染木薯‘JG1301’和‘SC9’叶片不同时间点cDNA等量混合作为模板,进行普通PCR扩增。利用PCR对内参基因引物特异性进行检测,在2%(w)琼脂糖凝胶电泳中,内参基因扩增产物在100~250 bp间,且目标条带单一,无非特异性条带和引物二聚体,扩增片段条带大小符合预期 (图1-A)。以Xpm侵染木薯‘JG1301’和‘SC9’叶片后不同时间点的cDNA为模板,经过qRT-PCR扩增后,对候选扩增片段的溶解曲线进行分析,结果表明,所选择的5个木薯候选内参基因引物特异性好,无杂峰,可用于后续内参基因稳定性研究(图1-B)。
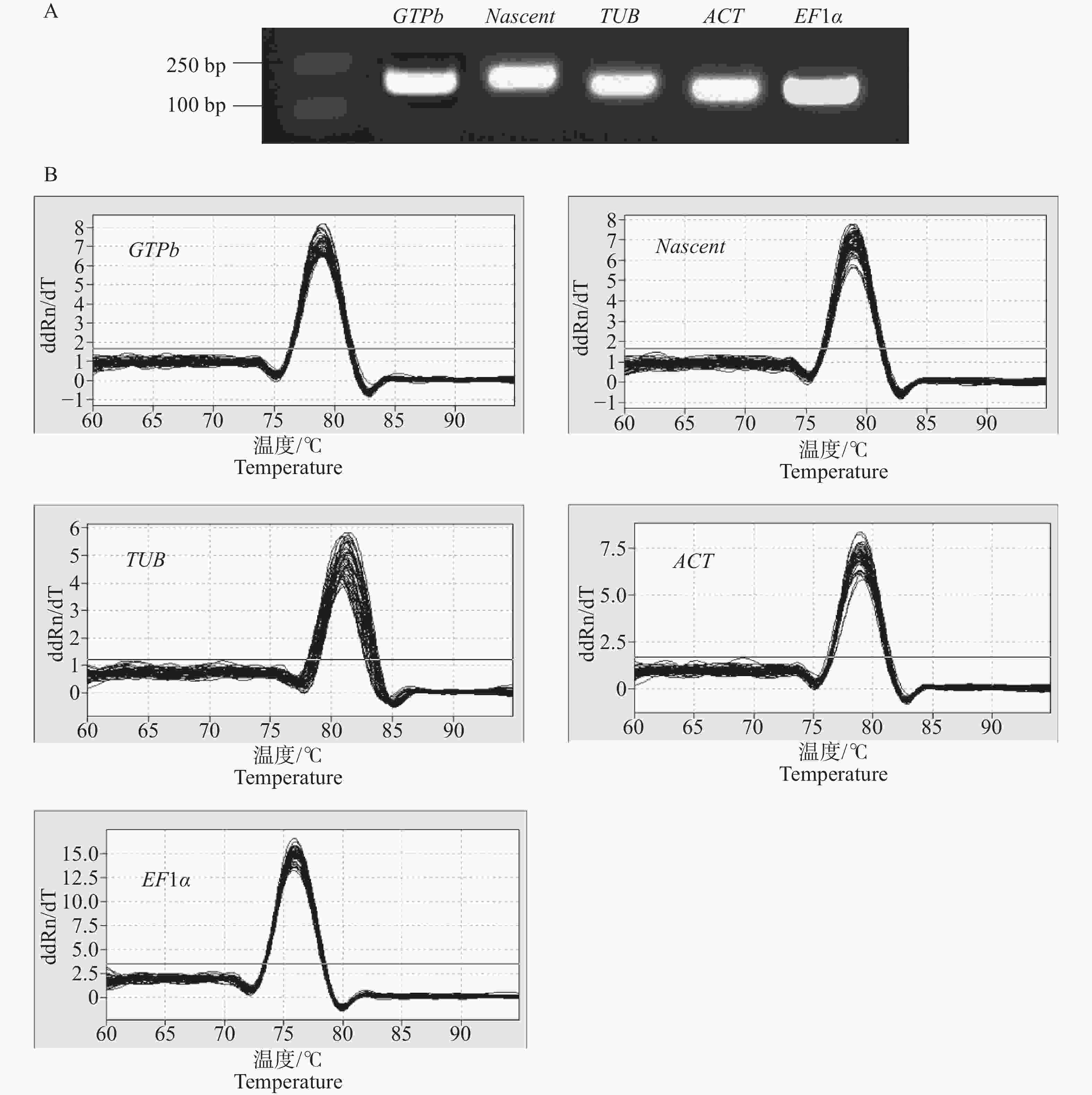
Figure 1. Reference gene primer detection
通过5个木薯候选内参基因引物的标准曲线得到引物的扩增效率和决定系数,结果表明,所选择的5个木薯候选内参基因荧光定量引物扩增效率在100.7%~109.4%,扩增效率最高的基因是EF1α,引物扩增效率为109.4%,引物扩增效率最低的是Nascent基因,扩增效率为100.7%。5个引物的决定系数R2 在0.993~0.999间,决定系数最高的基因是ACT基因,决定系数为0.999,决定系数最低的是GTPb基因,决定系数为0.993(表1)。
-
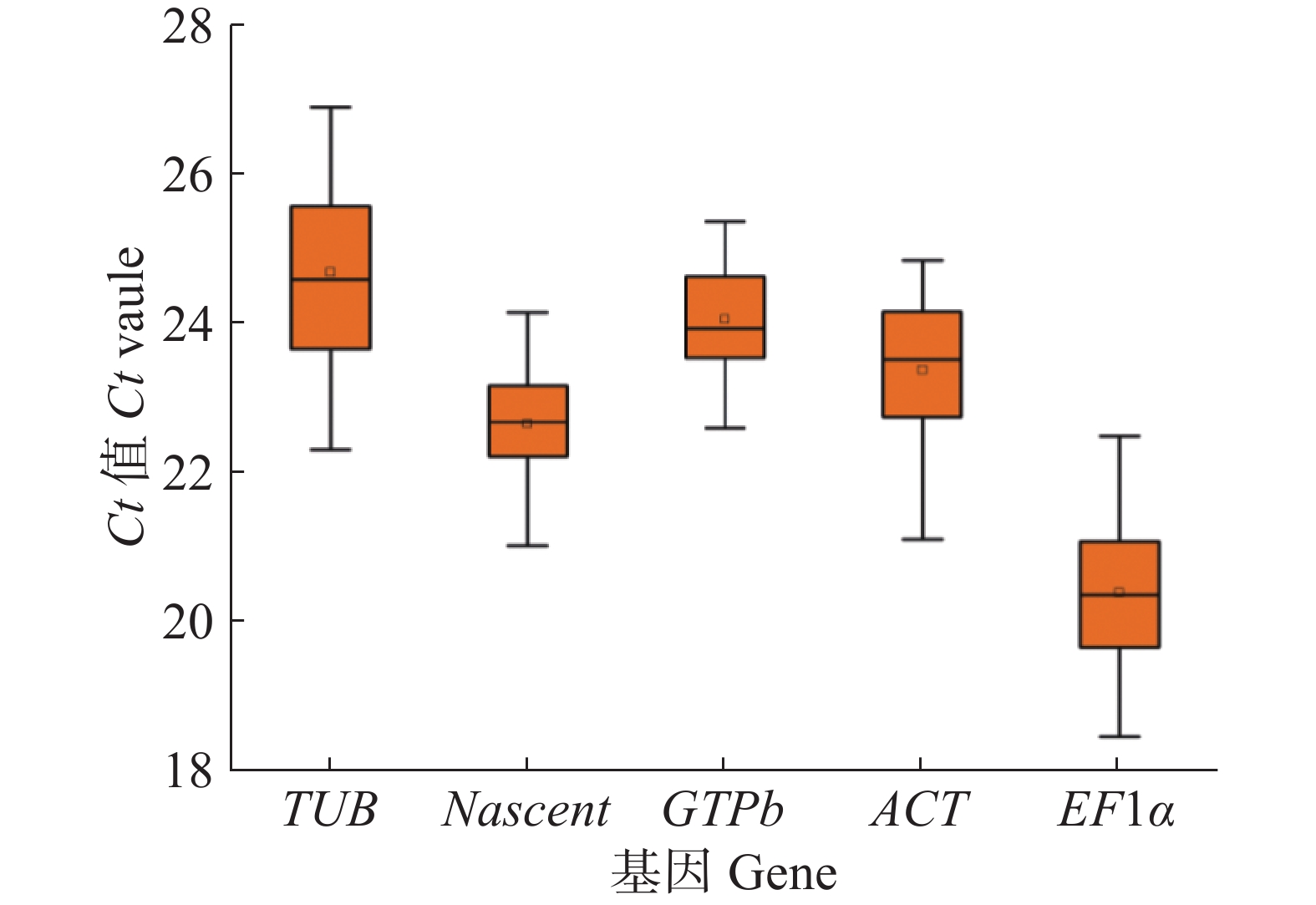
对 5 对木薯内参基因引物以Xpm侵染抗病品种‘JG1301’和感病品种‘SC9’木薯叶片后不同时间点的cDNA为模板进行qRT-PCR实验,通过检测其表达量得到对应的 Ct 值。Ct 值与基因的表达丰度成反比,Ct 值越大,基因的表达丰度越低,反之Ct 值越小,则基因的表达丰度越高。从图2可知,5个木薯候选内参基因中表达丰度最高的基因是EF1α,Ct 值在样本检测中为18.46~22.48;表达丰度最低的基因是TUB,Ct 值在样本检测中为22.30~26.89。
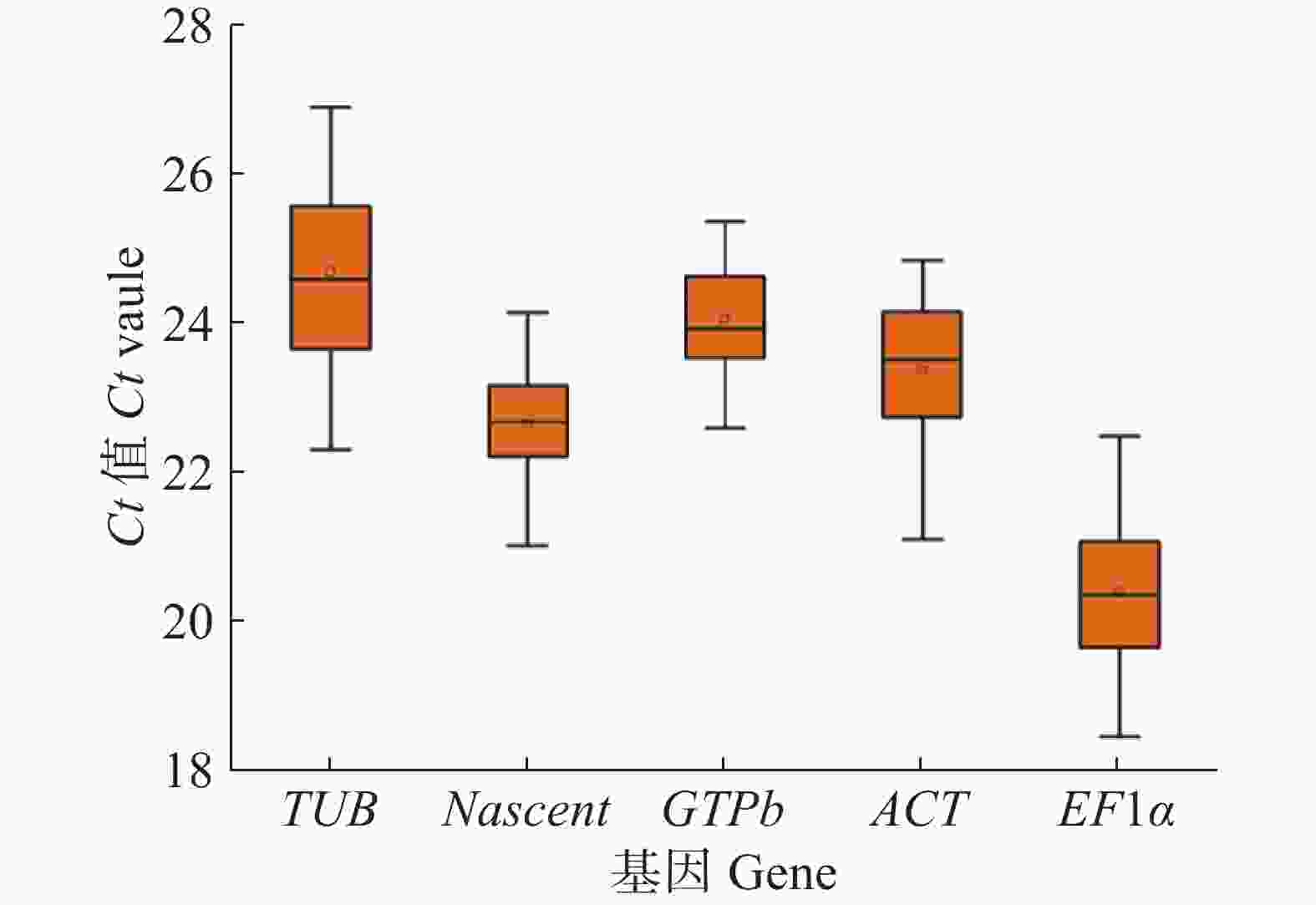
Figure 2. Ct value distribution of five candidate reference genes
-
Delta Ct 算法是用平均标准差(m SD)来评价内参基因的稳定性。m SD值的大小能够反映内参基因的稳定性,数值越小代表内参基因越稳定,越大则越不稳定。通过Delta Ct 方法计算5个木薯候选内参基因的稳定性可知,5个候选内参基因的m SD排序为TUB > EF1α > ACT > GTPb > Nascent, 其中m SD最大的是TUB,其m SD为1.18;m SD最小的是GTPb,其m SD为0.75,表明稳定性最好的是Nascent基因,稳定性最差的是TUB基因(表2)。
基因
Gene平均标准差
m SD排名
RankingNascent 0.75 1 GTPb 0.77 2 ACT 0.92 3 EF1α 1.00 4 TUB 1.18 5 Table 2. Results of Delta Ct Software
-
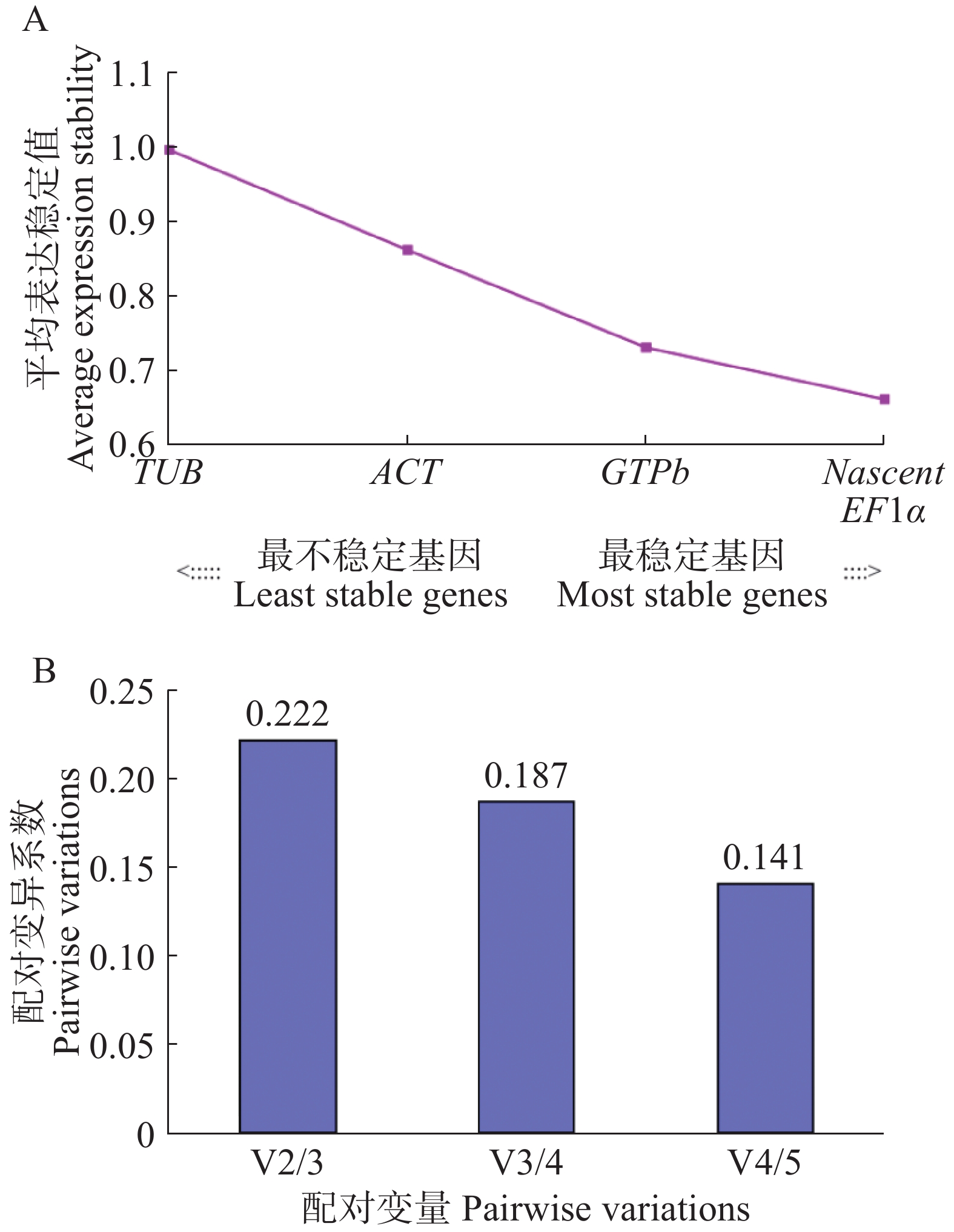
GeNorm 程序是根据平均变异度M值来衡量基因的稳定性,程序默认的取舍值为1.5,当M值<1.5 时,基因被认为相对稳定,M值越低则表示基因越稳定。该程序采用成对比较法(pairwise comparison approach)计算出基因表达稳定性的M值,根据Genorm默认变异系数(Vn/Vn+1)的值来确认最优的内参基因数目。当Vn/Vn+1<0.15时,则认为最佳的内参基因数为n个,若Vn/Vn+1>0.15还可引入1个新的内参基因后计算标准化因子的平均配对变异V值,并根据Vn/Vn+1 值来确定所需最适内参基因数目。
如图3所示,5个木薯内参候选基因的M值都小于1.5(图3-A),但是表达稳定性有较大差异,M值排序为TUB > ACT > GTPb >EF1α= Nascent,其中,M值最大的是ACT,M值为1.00,M值最小的是EF1α和Nascent,M值为0.66,说明稳定性最好的是EF1α和Nascent,稳定性最差的是TUB。其中,V4/5配对变异系数为0.141<0.15(图3-B),说明最适的内参基因数4个,不需要引入第5个基因进行校准。因此,本实验中最优基因数组合为4个,即EF1α、Nascent、GTPb和ACT。
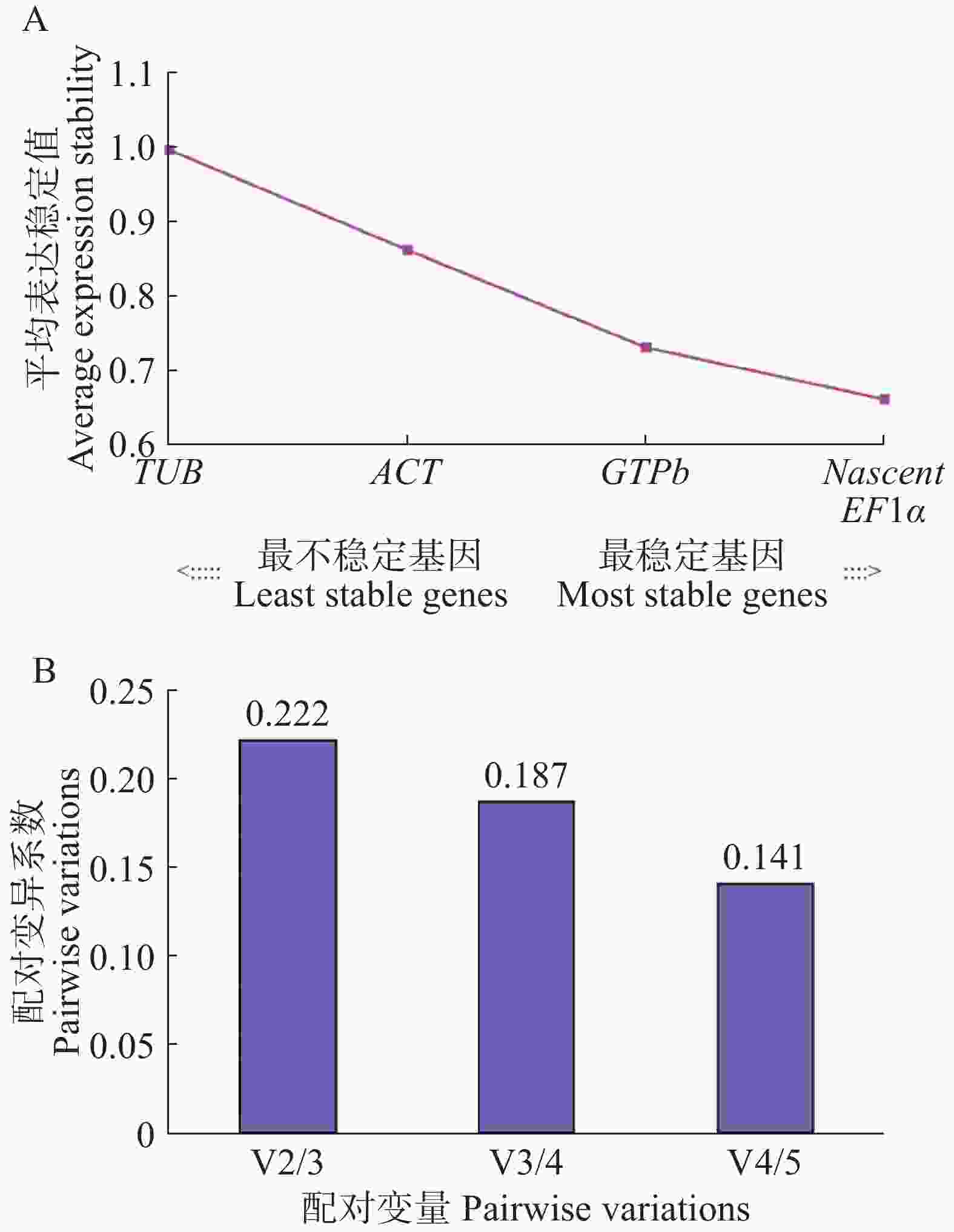
Figure 3. Genorm analysis
-
该程序基于模型分析,获得内参基因的表达稳定值,再根据稳定值大小来筛选出最合适的内参基因。NormFinder 程序根据候选内参基因的稳定值进行评价,稳定值越高表示稳定性越差,稳定值最小的基因,则为最稳定的基因。结果表明5个木薯候选内参基因的稳定值排序为TUB >ACT > Nascent > GTPb > EF1α,其中稳定值最大的是TUB,稳定值为0.714;稳定值最小的是EF1α,稳定值为0.348,说明稳定性最好的是EF1α基因,稳定性最差的是TUB基因(表3)。
基因
Gene稳定值
Stable value排名
RankingEF1α 0.348 1 GTPb 0.381 2 Nascent 0.444 3 ACT 0.520 4 TUB 0.714 5 Table 3. Results of Normfinder Software
-
BestKeeper 程序以内参基因 Ct 值的标准差(SD)来衡量基因的表达稳定性,SD值越小,表达越稳定,SD 临界值默认为1,当 SD值>1 时,说明该基因表达不稳定。采用重复配对相关分析和回归分析,通过计算Ct 值的标准偏差(SD)来确定稳定性较好的内参基因。结果表明,5个木薯候选内参的SD值均小于1,SD值排序为TUB > EF1α > ACT > GTPb > Nascent,其中SD值最大的是TUB,SD值为0.97;SD值最小的是Nascent,SD值为0.61,说明稳定性最好的是Nascent,稳定性最差的是TUB(表4)。
参数
Parameter基因 Gene TUB Nascent GTPb ACT EF1α 几何平均值[CP]
geo Mean [CP]24.65 22.64 24.044 23.36 20.37 算术平均值[CP]
ar Mean [CP]24.68 22.65 24.05 23.38 20.39 最小值[CP]
min [CP]22.30 21.02 22.59 21.1 18.46 最大值[CP]
max [CP]26.89 24.15 25.36 24.84 22.48 标准差[± CP]
SD [± CP]0.97 0.61 0.66 0.75 0.78 变异系数[% CP]
CV [% CP]3.91 2.68 2.73 3.21 3.84 调节系数标准差[± x-fold]
std dev [± x-fold]1.95 1.52 1.58 1.68 1.72 Table 4. Results of BesKeeper Software
-
RefFinder 在线软件可以对上述 4 种算法分析所得到的结果进行综合排序,赋予每个候选内参基因一个适当的权重,然后计算出其几何平均值,通过赋分综合排名,得分越低越稳定,筛选出表现最稳定的内参基因。结果显示RefFinder综合以上4种算法计算出来的稳定值排序为TUB > ACT > GTPb > Nascent > EF1α,其中稳定值最大的是TUB,稳定值为5.00;稳定值最小的是EF1α,稳定值为1.41,说明稳定性最好的是EF1α基因,稳定性最差的是TUB基因(表5)。
基因
Gene稳定值
Stable value排名
RankingEF1α 1.41 1 Nascent 1.73 2 GTPb 2.21 3 ACT 3.72 4 TUB 5.00 5 Table 5. Results of RefFinder Software
-
木薯种植和生产领域中,CBB是目前最为严重的木薯病害之一,根据种植环境条件的不同,引起的损失至高可达100%。因此,充分挖掘木薯抗病基因已成为木薯产业亟待解决的问题之一,而选择稳定的内参基因是木薯抗病基因挖掘的基础。木薯中稳定的内参基因筛选已有部分报道,如Moreno 等[19]研究了5对内参基因在木薯褐条病毒侵染木薯品种‘TMS60444’‘Ebwanateraka’和‘AR34’叶片中的稳定性,发现GTPb基因在感染木薯褐条病的条件下表达最稳定。Hu等[18]研究了26对内参基因在木薯‘Ku50’‘SC124’‘Rongyong9’‘SC5’和‘Arg7’等品种中不同发育时期的叶片、块根、叶柄及花及干旱胁迫下的不同时间点叶片合计21个样本中的内参稳定性,发现Nascent基因在绝大多数情况下都稳定表达[18]。在哥伦比亚Xpm生理小种UA681和CFBP1851侵染感病木薯品种
60444 后受诱导表达的木薯基因功能研究中,Carlos等[15]使用TUB基因作为内参基因,本研究所使用的Xpm生理小种 CHN 01 是在海南省分离得到的,将TUB作为候选内参基因研究Xpm 生理小种CHN 01侵染木薯下TUB在木薯中表达是否稳定[23]。ACT基因被认为是在拟南芥抗病研究中最稳定的内参基因[20],本研究选择了序列相似度最高的木薯直系同源基因(Manes.13G086400,ACT)作为候选内参基因。EF1α基因也被用于CBB抗病基因筛选的内参基因使用[21-22]。目前,木薯黄单胞菌侵染后的木薯内参基因稳定性研究较少,因此选择了上述5对文献报道的木薯基因GTPb、Nascent、TUB、ACT、EF1α作为后续内参基因。本研究采用了Delta Ct、Genorm、NormFinder、BestKeeper以及在线软件Reffinder对所选用的5对内参基因在木薯抗感品种(‘JG1301’和‘SC9’)不同接种时间点进行了稳定性打分,进一步筛选出在Xpm侵染木薯的条件下仍然能稳定表达的内参基因。结果表明,qRT-PCR数据显示5个候选内参基因都有不同幅度的变化,研究发现,Nascent基因在木薯不受外界胁迫的情况下可以稳定表达,但是在干旱胁迫下表达不稳定[18],结合Nascent基因在Xpm侵染木薯叶片后在木薯叶片中稳定性排名第2,说明外界生物与非生物胁迫影响了Nascent基因的表达量。GTPb基因在Xpm侵染木薯叶片后表达稳定性综合排名第3,而在木薯褐条病毒侵染木薯条件下较为稳定[19],可能因为褐条病毒侵染机理与Xpm作为维管束病害的侵染机制不同,造成了GTPb基因在不同生物胁迫条件下的表达量变化差异。
Carlos在木薯抗病基因研究中使用了TUB作为内参基因,同时Wei等[24]在进行木薯耐盐研究中也使用TUB作为内参基因。Tong等[25]研究发现,TUB基因在桃的不同组织中表达稳定,Zhong等[26]研究发现TUB基因在荔枝的不同组织和遮光处理下都能稳定表达,但是Nicot等[27]的研究表明,TUB在马铃薯侵染晚疫病和冷胁迫实验中表达量不稳定。本研究结果表明,TUB在Xpm侵染木薯后稳定性最差,说明TUB在木薯受到病原菌胁迫时表达不稳定。
ACT基因能在栾树的不同组织中稳定[28],ACT也能在百合花不同发育时期和不同组织中稳定表达[29],但Nicot等[27]研究发现,在马铃薯侵染晚疫病和盐处理下ACT基因在马铃薯叶片中表达最不稳定,Li等[30]在甘薯内参基因筛选实验中发现,在病毒感染和非病毒感染的条件下甘薯叶片中的ACT基因表达量不稳定。本研究发现,Xpm侵染木薯叶片后ACT基因在木薯叶片中表达不稳定,说明ACT基因虽然在多种植物中被广泛用作内参基因,但是不适合在部分薯类作物中当作内参基因。
Chang等[21]和Zheng等[22]在进行CBB研究时均使用EF1α作为内参基因;Nicot等[27]研究发现,在马铃薯晚疫病侵染下和冷胁迫下马铃薯叶片中表达最稳定的内参基因是EF1α。Mascia等[31]研究发现, EF1α基因在菊苣叶片和根组织中的表达稳定性最高。唐枝娟等[32]研究发现,强弱致病力的白叶枯病菌PXO99和PXO61侵染下的水稻EF1α基因表达最稳定。本研究结果也表明,在Xpm侵染下,木薯EF1α基因在抗感木薯不同时间点表达量最为稳定。综合上述结果可知,在多种植物和多种外界胁迫下,EF1α基因都能在植物叶片中稳定表达,可作为一个稳定的内参基因使用。本研究结果为木薯抗细菌性枯萎病相关抗病基因挖掘研究中稳定内参基因的使用奠定了理论基础
Screening and Evaluation of Reference Genes in Cassava unpon Infection by Xanthomonas phaseoli pv. manihotis
DOI: 10.15886/j.cnki.rdswxb.20240201
- Received Date: 2024-12-30
- Accepted Date: 2025-02-23
- Rev Recd Date: 2025-02-16
- Available Online: 2026-02-28
- Publish Date: 2026-03-25
-
Key words:
- cassava bacterial blight /
- Manihot esculenta /
- reference gene /
- qRT-PCR
Abstract: In order to screen for stable expression of cassava reference genes in cassava leaves infected with Xanthomonas phaseoli pv. manihotis (Xpm), we used Xpm resistant variety ‘G1301’ and susceptible variety ‘SC9’cassava leaves as research materials. Samples were collected from cassava leaves of ‘G1301’ and ‘SC9’ at 0 h, 6 h, 12 h, 24 h, 72 h, and 120 h after Xpm infection, and the stability of the commonly used reference genes Nascent, EF1a, ACT, GTPb, and TUB was determined. The results showed that there were differences in the expression levels of five reference genes after Xpm infection in the leaves of ‘G1301’ and ‘SC9’. Delta CT, GeNorm, NormFinder, BestKeeper and RefFinder software were used to rank the stability of these candidate reference genes. We found that the most stable reference gene expressed in Xpm-infected leaves is EF1α. This study identified the stable internal reference genes expressed during Xpm infection in cassava leaves, laying the foundation for studying gene expression in response to Xpm infection and exploring disease resistance genes.
| Citation: | Li Ciyun, Yang Jing, Lu Dongying, Yang Jianfei, Yang Qi, Wang Fei, Chen Yinhua, Niu Xiaolei. Screening and Evaluation of Reference Genes in Cassava unpon Infection by Xanthomonas phaseoli pv. manihotis[J]. Journal of Tropical Biology, 2026, 17(2): 215-222. doi: 10.15886/j.cnki.rdswxb.20240201

|
 DownLoad:
DownLoad: