
-
茉莉酸(jasmonic acid,JA)是植物体内重要的内源激素之一,其与衍生物茉莉酸甲酯MeJA、异亮氨酸偶联物JA-Ile等统称为茉莉酸类物质(jasmonates,JAs)。JAs在植物生长发育及抵御昆虫、食草动物取食、坏死性病原菌侵染等生物胁迫及干旱、盐害等非生物胁迫中发挥重要作用[1-2]。健康植株中JAs含量很少,JA应答基因被JA信号转导关键阻遏物JAZ(jasmonate ZIM domain)抑制。一旦植物受伤或遭遇生物非生物胁迫时,JA生物合成增加,进而形成JA-Ile等活性形式,COI1(coronatine-insensitive 1)受体复合物会感知JA-Ile等并促进JAZ的泛素化降解,缓解对下游相关转录因子的抑制,最终启动JA应答基因的表达。随着JA含量不断增加,JAZ基因的表达也会逐渐被JA激活从而导致JA应答基因再次被抑制。JAs的持续积累会抑制植物生长,因而,及时进行JAs的分解代谢,对平衡植物生长发育与防御反应过程显得尤为重要[3-4]。植物体内JA-Ile的含量直接影响JA信号的强度,控制JA-Ile的丰度和动态对于正确调节JA响应至关重要。JA-Ile的分解代谢主要通过ω氧化和解偶联2种途径实现,前者通过内质网定位的细胞色素P450氧化酶CYP94B1/B3将JA-Ile羟基化成12-OH-JA-Ile,进而通过CYP94C1氧化12-OH-JA-Ile形成12COOH-JA-Ile[5-6];后者依赖于酰胺水解酶IAR3和ILL6裂解JA-Ile形成JA,同时也可作用于12OH-JA-Ile形成无活性形式12-OH-JA[7-8]。然而拟南芥(Arabidopsis thaliana)iar3/ill6基因双突变体依然可以检测到12-OH-JA,意味着必然存在其他途径可以产生12-OH-JA[3]。
很长一段时间,人们都未在植物体内发现JA是如何直接羟基化成12-OH-JA的。有意思的是,来自稻瘟病菌(Magnaporthe oryzae)的单加氧酶Abm却可以直接将植物内源JA羟基化从而干扰水稻(Oryza sativa)免疫[9]。2017年,拟南芥茉莉酸氧化酶(jasmonate-induced oxygenases,JOXs/jasmonic acid oxidases,JAOs)JOXs蛋白被发现具有直接羟基化JA形成12-OH-JA的酶活性[3,10]。2021年,首次报道了拟南芥JOX2与底物JA、共底物2-酮戊二酸(2-oxoglutarate,2OG)和亚铁原子复合物的晶体结构,分析并验证了JOX2结合JA的分子机制[11]。JOXs蛋白属于植物基因组中第二大酶家族2-酮戊二酸依赖型双加氧酶2OGD(2-oxoglutarate-dependent dioxygenase)基因家族DOXC46分支类蛋白, 绝大部分2OGD蛋白含有DIOX-N (PF14226,Non-haem dioxygenases in morphine synthesis N-terminal)和2OG-Fe(Ⅱ)(PF03171,2OG-Fe(Ⅱ)oxygenase)结构域;DOXC类蛋白是植物中最大、功能最多样的一类加氧酶,参与植物激素如乙烯、赤霉素、生长素、水杨酸等合成或分解代谢过程,其中JOXs蛋白直接参与茉莉酸分解代谢[3,12]。拟南芥AtJOX1/2/3/4基因显著受到JA诱导表达,AtJOX1/2/4蛋白均能够单独羟基化JA,但AtJOX3的酶活性很弱;4种基因全突变体joxQ表现出JA信号通路升高相似的发育表型,植物明显比野生型弱小,但突变体中防御相关基因表达增加并且对坏死型病原真菌灰霉菌(Botrytis cinerea)和甘蓝夜蛾(Mamestra brassicae)幼虫抗性提高[3]。水稻中也存在类似的现象,OsJOX4酶活性也很弱,体外酶活实验几乎检测不到12-OH-JA产生,joxs全基因突变体水稻生长弱小,突变体中JA通路Marker基因的基础表达水平上升,对M. oryzae稻瘟病菌的抗性提高[13]。渐狭叶烟草(Nicotiana attenuata)中4个JOX-like类基因分别沉默的植株中均检测到了不同含量的12-OH-JA产生,但沉默JOX-like基因并不影响烟草生长表型,这可能与VIGS基因沉默的效果和时效性有关,同时沉默植株也增强了对斜纹夜蛾(Spodoptera litura)的抗性[14]。棉花(Gossypium hirsutum)GhJOX2基因在抗病品种棉花中表达量显著低于感病品种,GhJOX2沉默植株对黄萎病具有更高抵抗力[15]。热胁迫会影响植物生长发育,拟南芥AtJOX1/2/3/4和AtST2A磺基转移酶基因高温下表达增加,共同促进12HSO4-JA形成从而降低具有生物活性茉莉酸盐的水平,导致JAZ蛋白积累,促进植物在较高的温度下生长[16]。以上这些研究结果表明,植物JOXs基因参与JA信号通路的调控。
木薯(Manihot esculenta)属于大戟科(Euphorbiaceae)木薯属(Manihot)多年生直立灌木,生产上多为一年生栽培品种,与马铃薯(Solanum tuberosum)、甘薯(Dioscorea esculenta)并称世界三大薯类作物[17]。木薯耐旱耐贫瘠,粗生易长,能广泛适应多种土壤及气候条件。木薯块根淀粉含量丰富(占干物质含量85%以上),可直接煮熟食用或加工成木薯制品;木薯块根中支链淀粉含量也较其他薯类作物高,因而也常用于造纸业、纺织业及生物燃料生产等,是热区重要的粮食及经济作物[18-19]。自19世纪20年代引种以来,中国华南地区已有200余年木薯种植历史,随着木薯功能特性的不断深入开发利用,中国对木薯需求量与日俱增,已成为世界上最大的木薯进口国,研究和发展木薯产业符合中国生物能源与粮食生产和谐发展的需求。木薯生产中面临着多种环境及生物胁迫,茉莉酸氧化酶JOXs基因家族成员已有报道在拟南芥、烟草等模式植物应对逆境胁迫中发挥重要作用,但尚未在木薯中有相关报道。因此,本研究旨在鉴定木薯的JOXs基因家族,分析其基因结构、保守结构域、系统进化关系、组织表达特异性及其JA处理或病原菌侵染下的表达模式,为阐明木薯JOXs家族的基因功能及木薯抗逆品种选育奠定基础。
-
木薯AM560-2参考基因组v8.1及拟南芥TAIR10基因组下载于Phytozome v13.0数据库(
https://phytozome.jgi.doe.gov/ ),通过以下2种方法综合确定木薯MeJOXs家族基因。第1种是通过Pfam(http://pfam.xfam.org/ )数据库下载2-OGD家族基因的隐马可夫模型PF03171,使用HMMER v3.3.2[20]搜索木薯蛋白组(E-value<10-5),初步候选木薯2OGD家族成员;随后通过SMART(https:smart.embl.de/)、NCBI-CDD(https://www.ncbi.nlm.nih.gov/cdd/ )、Pfam等数据库检验候选蛋白是否至少含有1个2OG-Fe(Ⅱ)结构域;根据拟南芥2OGD家族基因氨基酸序列[12],利用MUSCLE v5.3[21]软件进行木薯及拟南芥2OGD蛋白氨基酸多序列比对,trimAl v1.4.1[22]修剪对齐序列,IQ-TREE v1.6.12[23]最大似然法构建系统进化树;依据拟南芥AtJOXs家族基因在进化树中位置筛选出木薯MeJOXs成员。第2种是利用拟南芥中已知的4个AtJOXs基因蛋白序列本地BLAST(E-value<10-10)搜索木薯蛋白组,初步候选木薯JOXs蛋白,进一步检验候选蛋白是否含有DIOX-N及2OG-Fe(Ⅱ)结构域,确定木薯MeJOXs家族成员。 -
ExPASy工具(
http://www.expasy.org/tools )计算MeJOXs蛋白的理化特性。DeepTMHMM-1.0工具(https://services.healthtech.dtu.dk/services/DeepTMHMM-1.0/ )分析MeJOXs跨膜结构域。WoLF PSORT在线工具(https://wolfpsort.hgc.jp/ )、Cell PLoc 2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/ )等综合预测MeJOXs亚细胞定位。 -
以木薯、拟南芥、水稻、烟草JOX蛋白氨基酸序列为基础,利用MUSCLE v5.3软件进行氨基酸多序列比对,随后利用MEGA 7.0进行NJ法(Neighbour-joining)构建系统发育进化树,设置p-distance模型,Bootstrap值设为
1 000 。 -
根据木薯基因组注释文件中MeJOXs家族成员基因结构信息,TBtools-Ⅱ v2.142绘制基因结构[24]。利用TBtools-Ⅱ从木薯基因组序列中提取MeJOXs家族成员转录起始位点上游
1500 bp序列,PlantCARE在线工具(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )分析启动子区顺式作用元件[25],TBtools-Ⅱ可视化顺式作用元件预测结果。利用MEME Suite 5.5.7在线工具(http://meme-suite.org/ )分析MeJOXs蛋白保守基序,motif最大数量设为10,其余参数保持默认值[26],TBtools-Ⅱ可视化motif预测结果。 -
木薯不同组织转录组数据(TCODP0016)下载于热带作物多组学数据库(
https://ngdc.cncb.ac.cn/tcod ),MeJA处理下木薯差异表达信息来源于文献记载[27];木薯细菌性枯萎病病原菌菜豆黄单胞菌木薯萎蔫致病变种Xpm(Xanthomonas phaseoli pv. manihotis)侵染木薯‘华南8号’(‘SC8’)种质转录组来自本实验室;利用TBtools-Ⅱ软件基于TPM(transcripts per million)值绘制基因表达热图,基于Log2 FC(log2 fold change)值绘制差异表达基因热图。 -
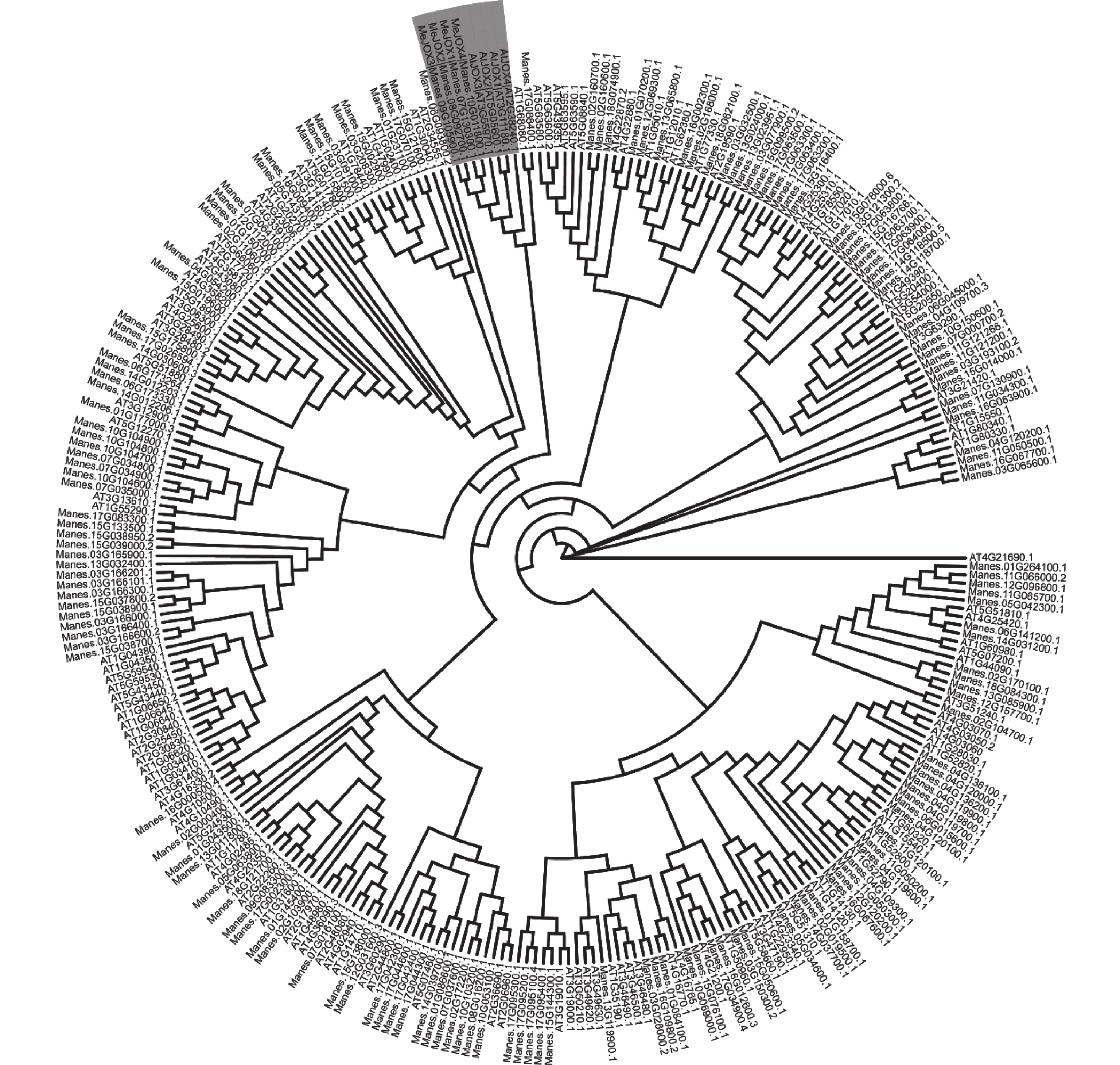
利用HMMER筛选木薯蛋白组,剔除不含特定结构域的蛋白后共获得165条候选的木薯Me2OGDs家族成员,随后与拟南芥130条At2OGDs成员构建最大似然树(图1)。由于拟南芥的AT1G68080.1基因虽然属于At2OGD家族但不属于任何一个亚族,尚未确定分组,且与其进化上最相近的木薯Manes.17G088400.1基因与同分支其余四个基因同源性极低,因而根据拟南芥AtJOXs基因(JOX1:AT3G11180.1,JOX2:AT5G05600.1,JOX3:AT3G55970.1,JOX4:AT2G38240.1)所在分支最终初步候选了4条木薯MeJOXs基因家族成员。
Figure 1. Phylogenetic analysis of 2OGD proteins identified in cassava and Arabidopsis
进一步利用拟南芥AtJOXs蛋白氨基酸序列与木薯蛋白组进行本地BLASTP比对,结合进化树分析结果共筛选出4条同时具有2OGD家族特异的DIOX-N和2OG-Fe(Ⅱ)结构域的木薯JOXs蛋白(图2),分别位于木薯第7、8、9、10号染色体,根据在染色体上的位置依次命名为MeJOX1~4。氨基酸多序列比对表明木薯MeJOXs蛋白与拟南芥AtJOXs蛋白相似度均在60%左右,MeJOXs蛋白成员间至少具有68%同源性,其中MeJOX1与MeJOX4、MeJOX2与MeJOX3两两之间最为相似,蛋白同源性分别达到83%、88%。木薯MeJOXs蛋白的氨基酸数目基本一致,除MeJOX3含有362个氨基酸外,其余均含有364个氨基酸;蛋白分子质量大小为40.9~41.2 kDa;理论等电点位于5.68~6.96,属于偏酸性蛋白;蛋白亲水性平均系数均小于0,表明4个木薯MeJOXs蛋白均为亲水性蛋白。DeepTMHMM均未预测到MeJOXs蛋白含有跨膜结构域,亚细胞定位预测结果表明所有家族成员均最有可能定位在细胞质中,可能在细胞质中发挥功能(表1)。

Figure 2. Multiple sequence alignment of JOX proteins in cassava and Arabidopsis
名称
Name基因号
Gene ID氨基酸数
Number of
amino acid分子质量/kDa
Molecular
weight理论等电点
Theoretical pI亲水性平均系数
Grand average of
hydropathicity亚细胞定位
Subcellular
localizationMeJOX1 Manes.07G130300.1 364 41.2 6.47 −0.419 细胞质
CytoplasmMeJOX2 Manes.08G092700.1 364 41.2 6.41 −0.364 细胞质
CytoplasmMeJOX3 Manes.09G105900.1 362 41.1 6.96 −0.346 细胞质
CytoplasmMeJOX4 Manes.10G011800.1 364 40.9 5.68 −0.287 细胞质
CytoplasmTable 1. Physical and chemical properties of cassava JOX proteins
-
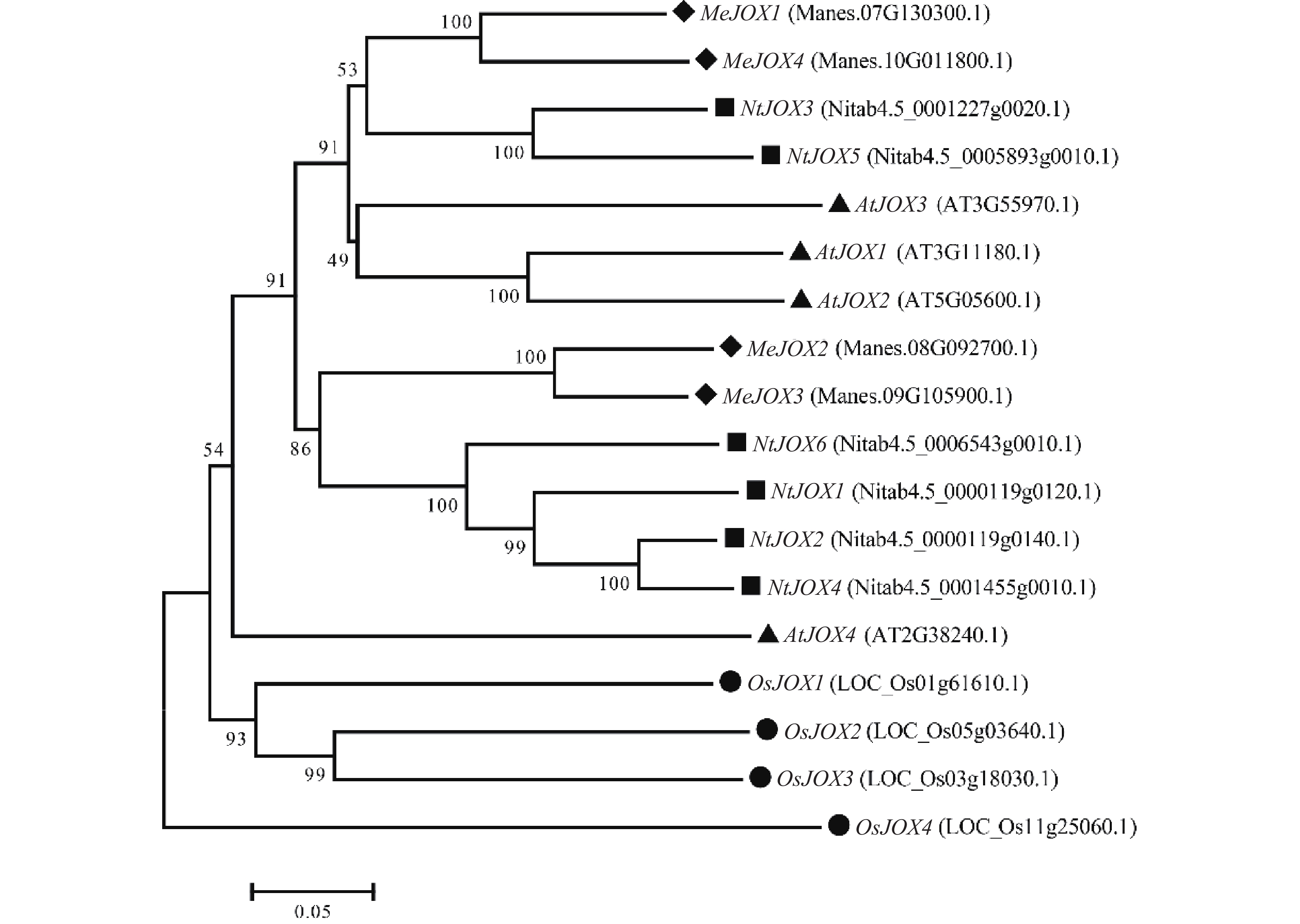
为了研究JOXs基因在不同物种间的进化关系,利用MUSCLE对拟南芥4个AtJOXs、水稻4个OsJOXs、烟草6个NtJOXs及木薯4个MeJOXs进行氨基酸多序列比对,随后利用MEGA7.0通过NJ法构建系统发育树(图3)。结果表明,同为双子叶植物的JOXs基因之间亲缘关系更近,木薯、拟南芥、烟草的JOXs基因间亲缘关系较水稻更近,水稻OsJOXs基因单独聚成一个分支。此外,木薯MeJOXs基因明显分为两个类群,MeJOX1/4基因聚为一类,MeJOX2/3基因聚为另一类,同一分支的MeJOXs基因间生物学功能可能类似;MeJOX1/4基因与拟南芥AtJOX1/2/3亲缘关系较近,预示着可能存在相似的生物学功能。
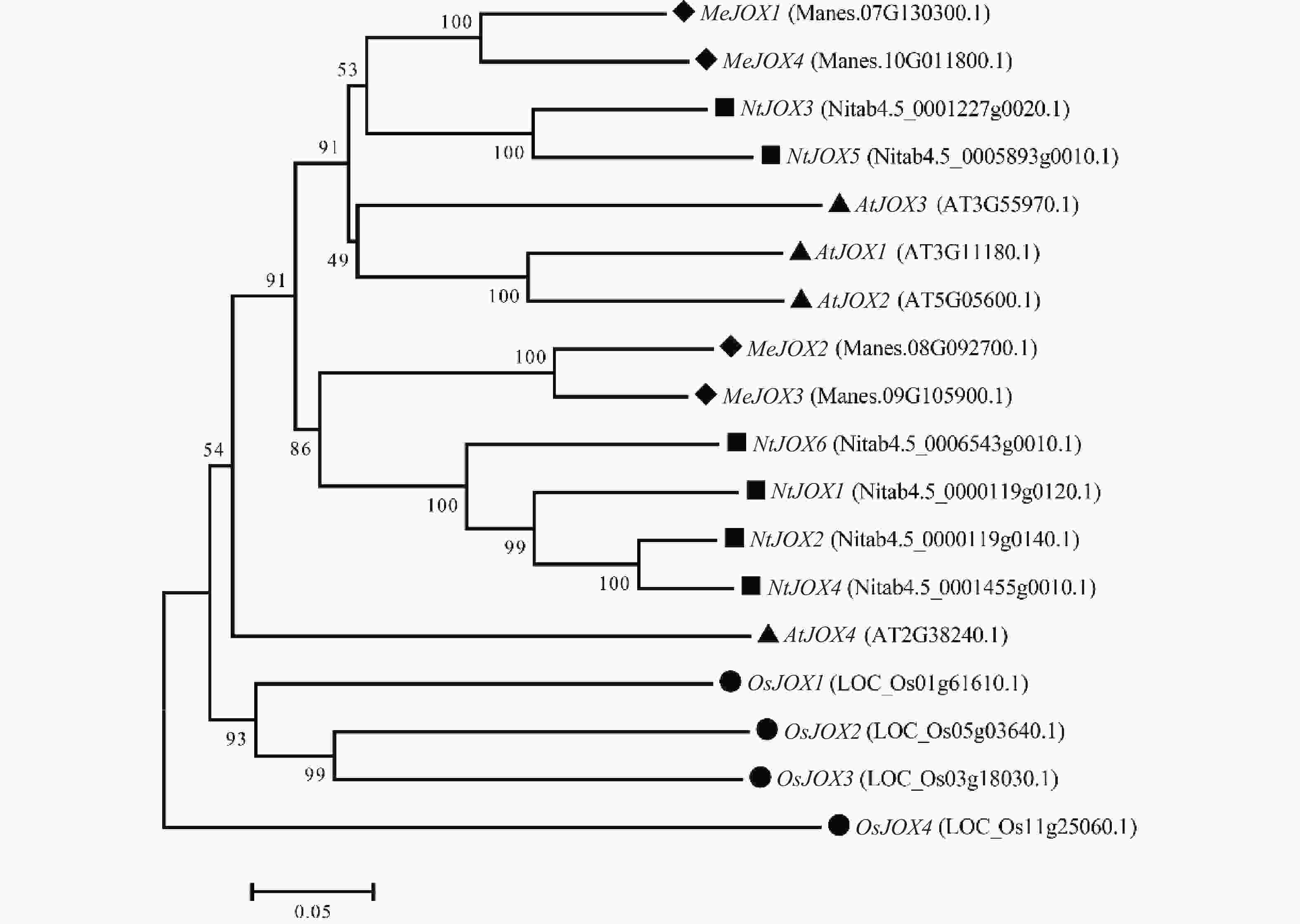
Figure 3. Phylogenetic tree of JOX genes in cassava, Arabidopsis, rice, and tobacco
-
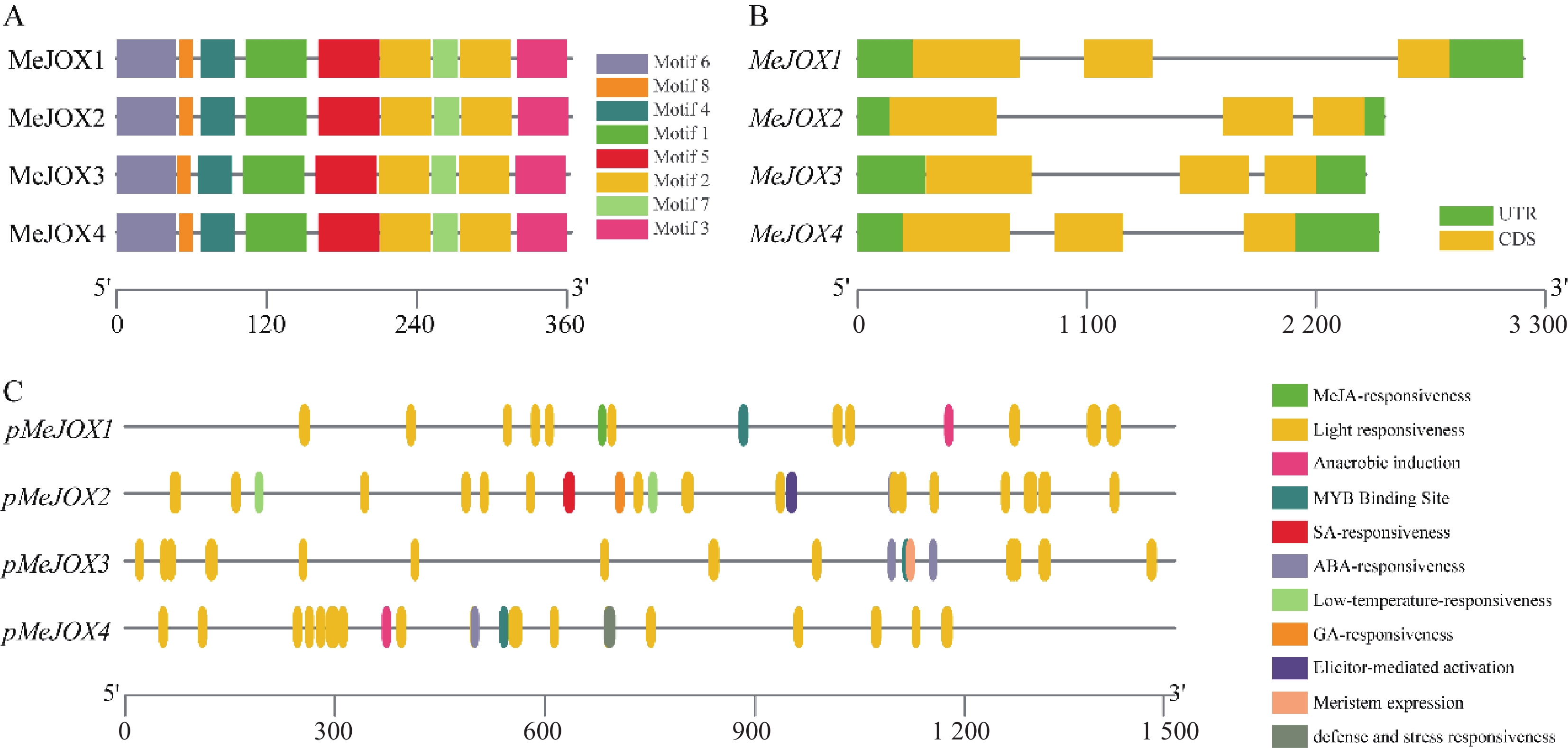
通过MEME对MeJOXs家族蛋白分析共发现8个保守的motif基序,每个成员间含有相似的motif分布(图4-A)。通过对MeJOXs家族成员基因结构分析发现,各个成员均包含5个外显子,2个内含子,其中3个外显子编码CDS区,其余2个为5´UTR和3´UTR非翻译区。此外,进化树中聚到同一类群的MeJOXs基因显示出相似的基因结构, MeJOX2和MeJOX3都表现出第1个内含子较长、第2个内含子较短的基因结构,而MeJOX1和MeJOX4第1个内含子较短,第2个内含子均较长(图4-B)。
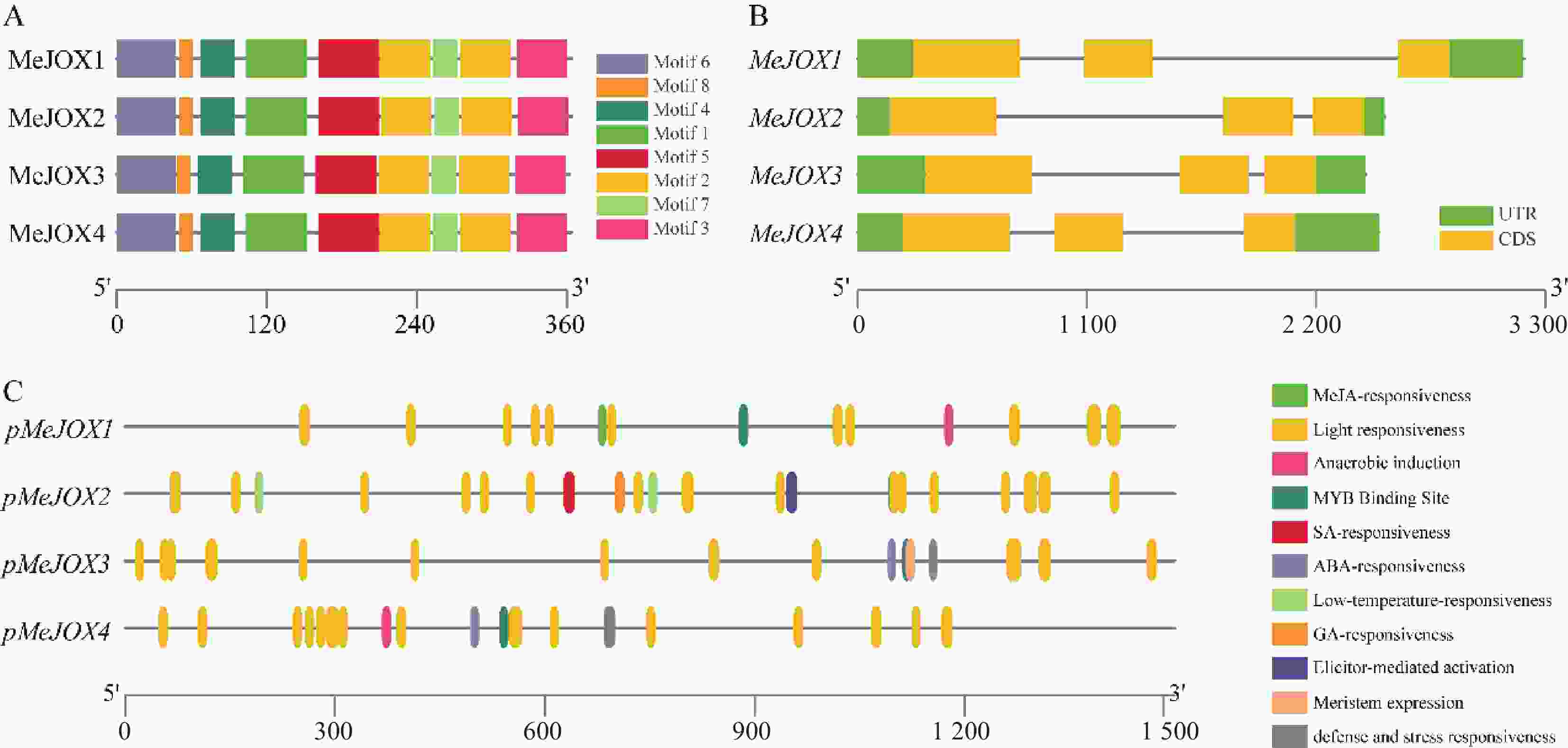
Figure 4. Gene structures, conserved protein motifs and cis-acting elements in the promoter regions of cassava MeJOXs members
通过对MeJOXs家族成员转录起始位点上游1 500 bp左右启动子区分析发现其启动子区包含多种不同类型的顺式作用元件,除了常见的CAAT-box、TATA-box等核心顺式作用元件外,还包含多种功能响应元件,例如光响应、激素响应、低温响应、厌氧响应元件、MYB结合位点等。每个MeJOXs成员启动子上分布最多的是光响应元件,包括AT1-motif、GA-motif、Box4、AE-box、TCT-motif、G-box、ACE、I-box、TCCC-motif等多种不同类型,表明MeJOXs可能响应昼夜光照变化;激素响应元件每个成员间分布不同,MeJOX1启动子含有2个TGACG-motif、CGTCA-motif类型的茉莉酸响应元件,MeJOX2启动子含有1个TCA-element类型的水杨酸响应元件、4个ABRE类型的脱落酸响应元件及1个P-box类型的赤霉素响应元件,MeJOX3和MeJOX4分别含有4个及1个ABRE类型的脱落酸响应元件;除此之外各个成员还含有自身特有的一些响应元件,如MeJOX1启动子中含有1个ARE类型的厌氧响应元件,MeJOX2启动子中含有2个LTR类型的低温响应元件、MeJOX4启动子中含有TC-rich repeat类型防御与胁迫相关响应元件(图4-C)。这些顺式作用元件的存在暗示MeJOXs基因可能在激素调节、环境胁迫等相应过程中发挥重要作用。
-
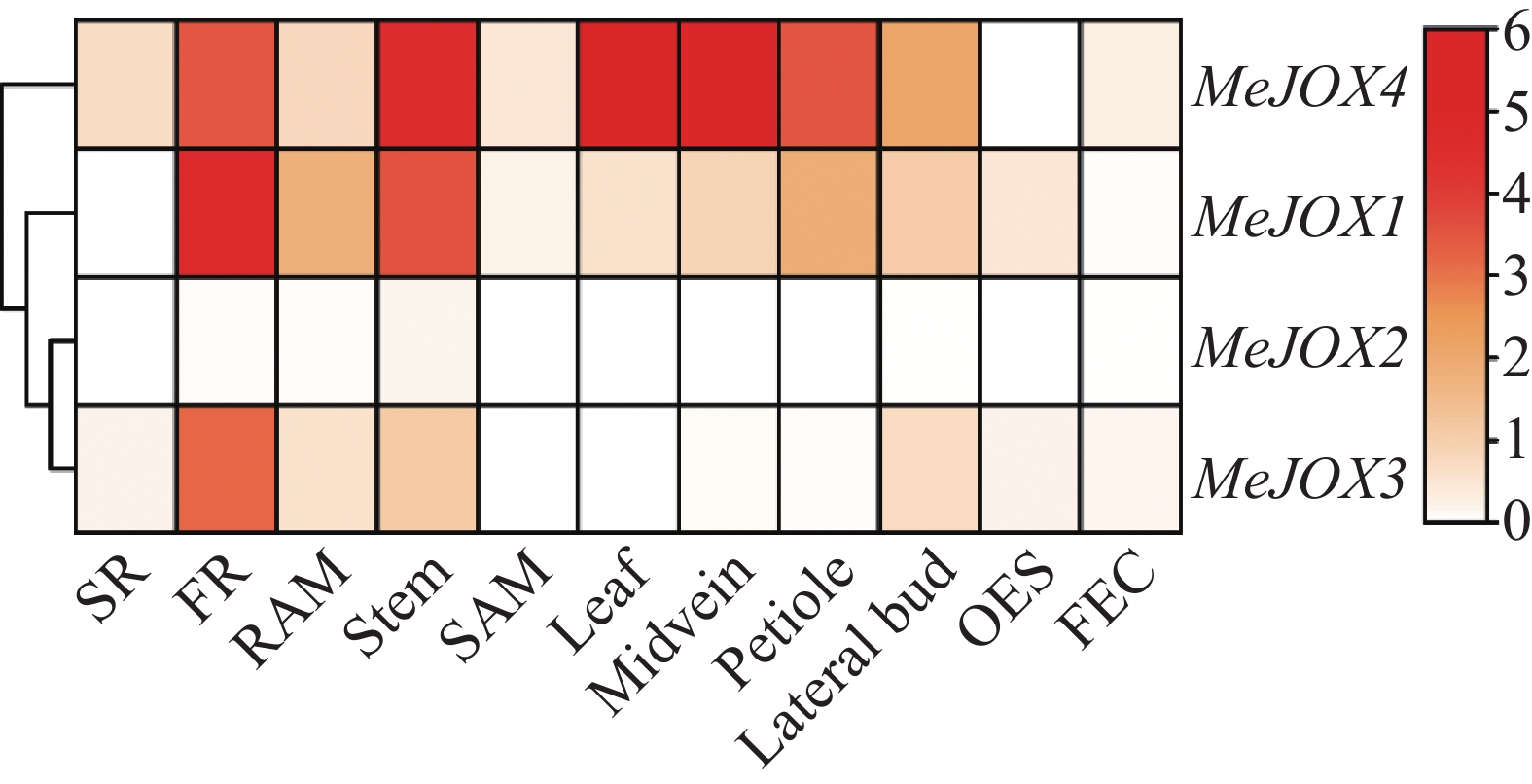
为了探究木薯MeJOXs家族基因的表达是否存在组织特异性,本研究利用木薯11种不同组织部位(块根、须根、根尖分生组织、茎、茎尖分生组织、叶片、叶中脉、叶柄、腋芽、体胚、脆性胚性愈伤组织)的公开转录组对MeJOXs的表达量TPM值进行log2(TPM+1)对数转换后绘制热图(图5)。结果显示,除MeJOX2在11种木薯组织中几乎均不表达外,其余3个MeJOXs家族成员在不同组织中具有不同的表达模式, MeJOX1在须根中显著高表达,MeJOX4在茎、叶片及叶中脉中均显著高表达,在须根、叶柄及腋芽中也有表达,MeJOX3在须根中有表达,但不如前两者。以上结果表明,在木薯不同组织生长发育过程中MeJOXs家族不同成员可能发挥不同的作用。
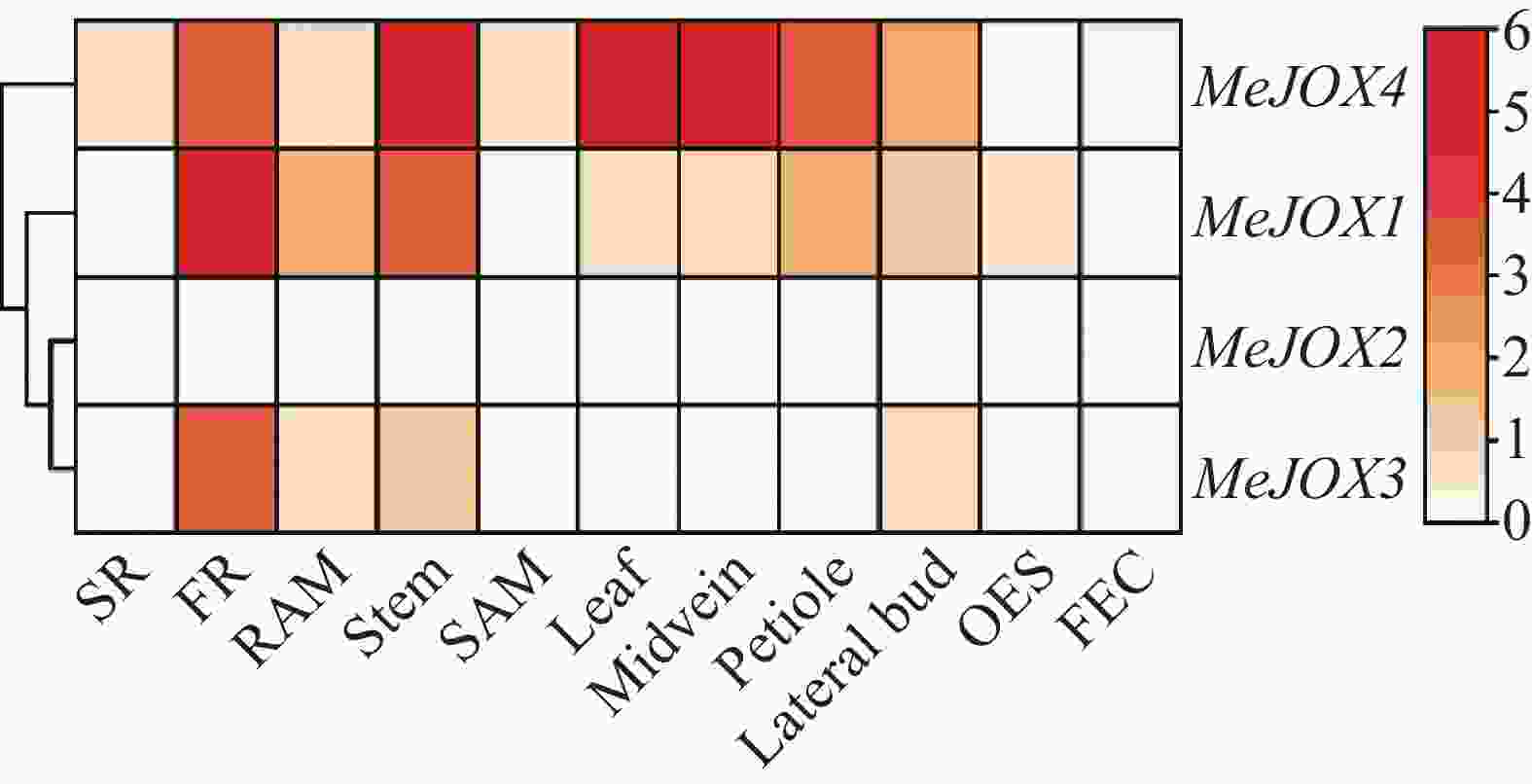
Figure 5. The expression heatmap of MeJOXs genes in different cassava tissues
-
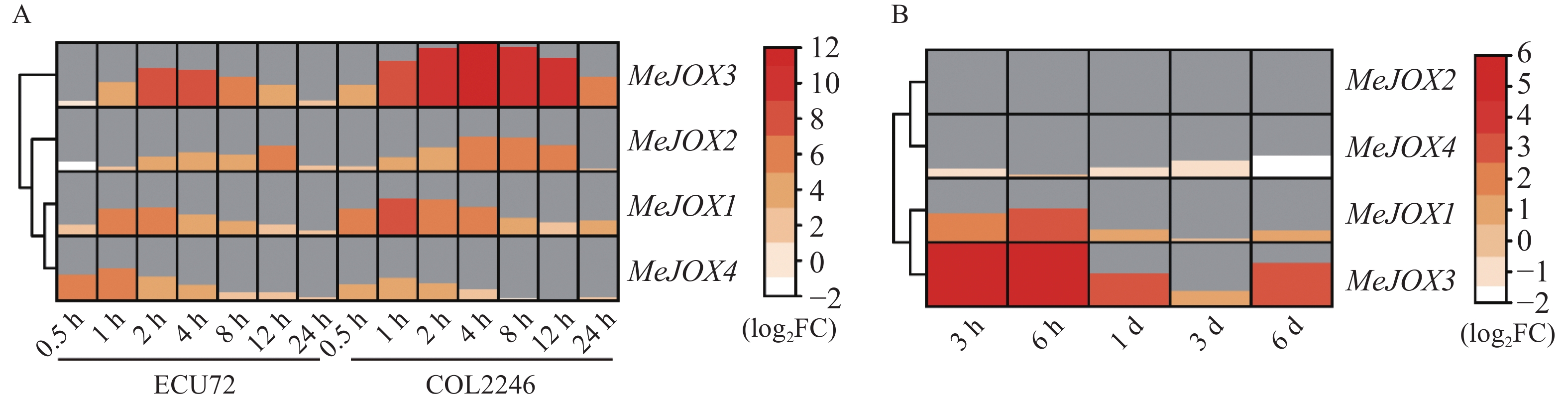
由于拟南芥AtJOXs基因均显著受到植物激素MeJA诱导表达,因而利用MeJA处理木薯种质ECU72、 COL2246的转录组分析了木薯MeJOXs家族基因在MeJA处理下的差异表达情况。结果如图6-A所示,在MeJA处理后木薯MeJOXs家族基因均能够响应且不同的时间点存在较为明显的差异表达(|log2 FC| ≥ 1且FDR≤0.05),MeJOX1/4层次聚类为一类,在2种木薯种质中表达模式更为接近,相对于MeJOX2/3,前两者在2种种质中MeJA处理0.5 h后即有明显上调表达趋势,处理后1~2 h表达达到峰值,随后表达逐渐下降;后两者在ECU72种质中,MeJA处理0.5 h后二者表达甚至稍呈下调趋势(log2 FC = −1.2或−0.6),随后表达才逐渐上调,2~4 h达到峰值后逐渐降低,但MeJOX2在MeJA处理12 h后表达水平还存在一个峰值起伏(log2 FC = 5.0)。同一层次聚类分支的基因之间表达模式也存在一些区别,例如MeJOX1在COL2246种质中受MeJA诱导表达幅度更大且更早达到峰值(1 h,log2 FC = 7.0),而MeJOX4在ECU72种质中受诱导表达幅度更大且更早达到峰值(1 h,log2 FC = 6.2)。除此之外,MeJOX2/3虽然在对应种质中表达模式较为相似,但MeJOX3在MeJA处理后多个时间点表达量上升幅度明显高于其他3个MeJOXs基因,且在COL2246种质中MeJA处理4 h后表达受诱导最为明显(log2 FC = 11.8)。以上结果说明,木薯MeJOXs家族基因受植物激素JA诱导表达。
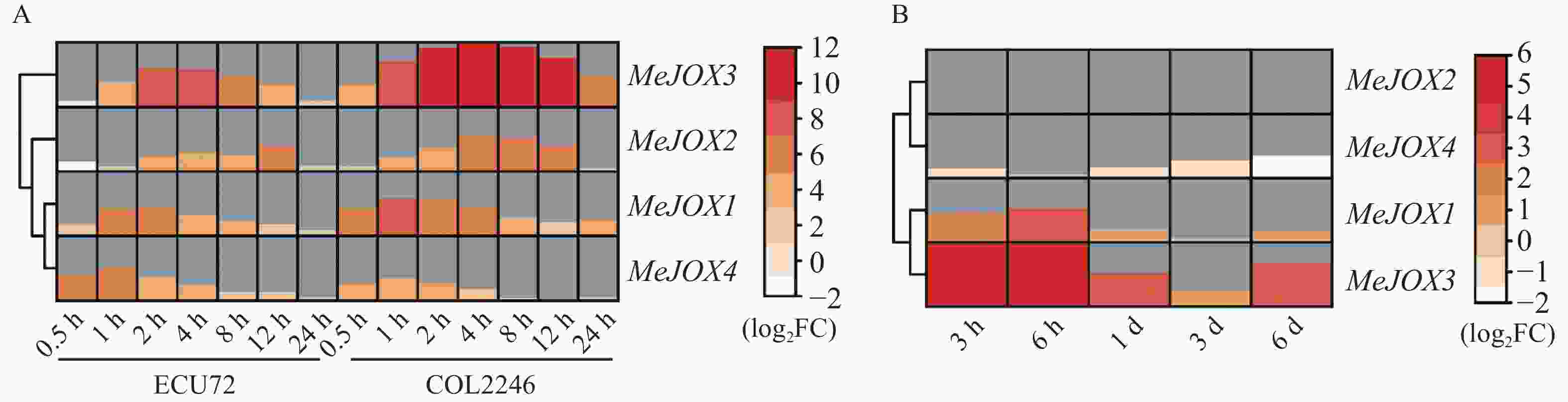
Figure 6. Heatmaps displaying differentially expressed genes of cassava MeJOXs under JA or pathogen Xpm treatments
已有多篇文献报道植物JOXs基因在病原菌侵染后表达量发生明显变化,因而利用本实验室已有木薯细菌性枯萎病菌Xpm侵染木薯‘SC8’种质后的转录组分析了MeJOXs家族基因响应病原菌Xpm侵染后的差异表达情况。结果如图6-B所示,除MeJOX2在病原菌侵染后各个时间点几乎不表达外(TPM≈0),其余3个MeJOXs基因在各个时间点均有不同程度差异表达(|log2FC| ≥ 1且FDR≤0.05),且响应Xpm侵染时MeJOX1/3、MeJOX2/4分别层次聚类为一支,表达模式相近。MeJOX1/3在病原菌侵染后表达即呈上调趋势,且MeJOX3受诱导表达情况更明显,病原菌侵染3 h后上调幅度即达到峰值(log2 FC = 5.2),其可能在病原菌侵染早期发挥生物学功能;而MeJOX4在病原菌侵染后呈下调表达趋势,6 d后达到峰值(log2 FC = −1.8),暗示其可能在病原菌侵染中发挥与MeJOX1/3相反的功能。以上结果表明,木薯MeJOXs家族基因多数都能响应病原菌Xpm侵染,且不同基因间生物学功能可能存在一定的差异。
-
植物2OGDs基因家族在激素合成和分解代谢过程中起着关键作用,2OGD家族分为DOXA、B、C 3个亚族,目前只有部分基因得到功能验证。茉莉酸氧化酶JOXs家族基因属于DOXC46分支基因,参与茉莉酸分解代谢过程,直接将JA羟基化为12-OH-JA,抑制了JA向JA-Ile活性形式转变的过程,对调节植物生长发育及应对生物和非生物胁迫至关重要。2OGDs基因家族目前已在多个植物中具有相关报道,例如拟南芥和水稻中分别含有130和114个[12],烟草中含有126个[28],然而木薯中尚未有2OGDs家族基因相关报道。本研究利用生物信息学手段初步鉴定到木薯2OGDs家族基因165个,多于以上几个物种,但JOXs亚族基因在每个物种中的数量变化不大,拟南芥和水稻中均只含有4个JOXs基因,烟草中有6个JOXs基因,木薯中也仅鉴定到4个MeJOXs成员,定位在相邻的4条染色体上。4个MeJOXs在基因结构方面较为类似,内含子外显子数量较少,可能与其需要快速激活应对外界胁迫有关[29]。此外,蛋白理化性质方面差异不大,4个MeJOXs蛋白含有类似的保守基序,且保守基序在蛋白上的分布情况基本一致,每个MeJOX蛋白N端和C端均分别含有保守的DIOX-N和2OG-Fe(Ⅱ)结构域,2OG-Fe(Ⅱ)结构域中负责结合Fe(Ⅱ)离子的基序HxDx…H和RxS基序在不同物种中也十分保守,基本不发生变化,暗示其功能在不同物种中具有保守性。JOXs蛋白具有直接羟基化JA形成12-OH-JA的酶活性,但拟南芥和水稻JOXs家族成员虽然多数成员均能够单独羟基化JA,但也都存在个别成员酶活性很弱[3,13],木薯中4个MeJOXs蛋白是否都具有较强的羟基化JA酶活还需进一步的体外酶活实验验证。
基因的表达具有时空特异性。木薯MeJOXs基因在不同组织中的表达存在明显差异,MeJOX2基因在各个组织中基本不表达,可能需要某些特定的生物或非生物胁迫才能刺激其表达,其余基因在某些特定的组织中显著高表达。例如MeJOX1在须根中显著高表达,可能与该组织的生长发育密切相关;对MeJOXs基因启动子区顺式作用元件预测结果表明每个成员启动子区均分布着大量不同类型的光响应元件,可能与光照条件可以影响茉莉酸的生物合成与降解有关,例如某些情况下强光可能促进茉莉酸的积累,弱光可能促进其降解[30],通过感知光照强弱变化调节MeJOXs的表达可能是植物调节茉莉酸动态平衡的关键;此外MeJOXs启动子区含有数量不等的茉莉酸、水杨酸、脱落酸等激素响应元件,说明MeJOXs基因的表达不光可能受茉莉酸调控,很可能还受其他植物激素的调控,可以更灵活地调节基因表达以适应不同的环境压力和信号整合。据报道拟南芥中4个AtJOXs基因均强烈受到MeJA诱导表达,尤其AtJOX1/2的表达量上调了400倍以上[3];水稻中除OsJOX4不受MeJA诱导表达外,OsJOX1/2/3也不同程度受MeJA诱导表达[13],OsJOX2最高上调表达约16倍,与二者类似,木薯MeJOXs基因也受到MeJA诱导表达,且在MeJA处理不同的时间点及不同的木薯种质中受诱导程度存在明显差异,MeJOX3受诱导表达最明显,最高时上调表达11.8倍,表明MeJOXs家族基因参与JA调控通路。
拟南芥中joxQ全基因突变体对灰霉菌的抗性增加[3],水稻中osjoxs突变体JA通路Marker基因的基础表达水平上升,并且对稻瘟病菌的抗性提高[13],这表明在拟南芥和水稻面对灰霉菌和稻瘟病菌侵染时JOXs基因负调控抗病过程。对木薯‘SC8’在病原菌Xpm侵染下转录组数据分析表明,除MeJOX2几乎不响应外,其余3个MeJOXs均能够响应病原菌入侵,但三者之间表达模式存在差异。MeJOX1和MeJOX3表达情况类似,在病原菌入侵早期即有明显响应,但MeJOX3响应程度更高,各个时间点诱导表达更明显,而MeJOX4响应病原菌侵染的表达模式明显异于MeJOX1/3,在病原菌侵染的多数时间点被抑制表达,推测三者之间功能可能存在差异,对病原菌侵染存在正调控和负调控机制。
本研究共鉴定到木薯4个MeJOXs基因,MeJOXs在进化上相对保守,在不同组织中的表达具有组织特异性,受JA诱导表达且存在品种差异,同时多数成员能够响应病原菌侵染。本研究为进一步研究木薯MeJOXs基因家族在木薯生长发育以及生物和非生物胁迫中的功能提供了理论基础。
Genome-wide identification and expression analysis of jasmonate-induced oxygenases JOXs gene family in Manihot esculenta
DOI: 10.15886/j.cnki.rdswxb.20240199
- Received Date: 2024-12-25
- Accepted Date: 2025-01-24
- Rev Recd Date: 2025-01-18
- Available Online: 2026-01-15
- Publish Date: 2026-01-25
-
Key words:
- cassava /
- jasmonate-induced oxygenases JOXs /
- gene family /
- expression analysis
Abstract: To explore the role of jasmonate-induced oxygenases JOX family genes in the growth, development, and stress resistance of cassava (Manihot esculenta), MeJOXs family members were identified in cassava genome by bioinformatics methods, and their gene structure, promoter cis-acting elements, evolutionary relationships and expression patterns were analyzed. The results showed that there were four MeJOXs family genes in cassava genome, and each member had similar gene structure, conserved motifs, and protein domains. Each gene member exhibited at least 68% protein homology, with the highest similarity observed between MeJOX1 and MeJOX4, as well as between MeJOX2 and MeJOX3. Promoter cis-acting elements analysis showed that MeJOXs contained numerous light-responsive elements. Furthermore, each gene member possessed a varying number of hormone-responsive elements. Phylogenetic analysis showed that MeJOXs were more closely related to JOX genes in dicotyledonous plants. Transcriptome analysis revealed that all the genes were differentially expressed in stems, leaves, midveins, and fibrous roots, with the exception of MeJOX2 that was scarcely expressed in various cassava tissues. MeJOXs were induced by MeJA in different cassava germplasm, with MeJOX3 demonstrating the most significant expression by inducing. Upon infection by pathogen Xpm, MeJOX1/3/4 responded promptly, but their response patterns were distinctly different. MeJOX1/3 were upregulated, whereas MeJOX4 exhibited a trend of downregulation, and MeJOX2 exhibited negligible response. This study provides a theoretical foundation for further elucidating the functions of the MeJOXs gene family in cassava.
| Citation: | Hu Wencheng, Zhu Shousong, Wang Yixuan, Yang Da, Chen Yinhua. Genome-wide identification and expression analysis of jasmonate-induced oxygenases JOXs gene family in Manihot esculenta[J]. Journal of Tropical Biology, 2026, 17(1): 29-38. doi: 10.15886/j.cnki.rdswxb.20240199

|
 DownLoad:
DownLoad: