
-
辣椒疫霉(Phytophthora capsici)是一种极具破坏性的作物卵菌,对辣椒种植造成巨大的经济损失[1]。辣椒疫霉寄主广泛、传播迅速、易产生抗/耐药性[2-4],以及分泌大量酶或效应蛋白促进病原菌侵染致病,造成严重的辣椒疫病防控障碍。内质网是真核细胞分泌蛋白与膜蛋白合成加工的重要场所。已有报道显示,病原菌分泌蛋白(包括效应子)经内质网加工与修饰对病原菌的致病力至关重要[5-7]。Sec62作为易位复合体的重要成员,在促进蛋白质前体向内质网转运过程中起关键作用[8-10]。它通过末端EF基序与Sec61互作,形成V型结构调控Sec61蛋白通道,Sec62缺失后引起Sec61蛋白通道变窄处于非活动状态,导致细胞对内质网应激敏感性增加[11-14]。近期研究发现,Sec62可作为内质网自噬受体,在内质网应激的恢复阶段被激活,参与组织生长发育和内质网稳态调节[15-16]。AtSec62在拟南芥中参与调控植物的生长发育,Atsec62缺失后导致植物生长发育延缓、花粉形态异常,且育性降低;Atsec62过表达会增强植物对内质网应激的耐受性[17-18];稻瘟菌MoSec62是酵母内质网膜转运蛋白Sec62p的同源蛋白,MoSec62缺失后稻瘟菌突变菌株M1575致病力完全丧失,同时MoSec62也可以回补酿酒酵母sec62缺失导致的温度敏感性[19]。然而,辣椒疫霉中尚未见PcSec62的相关研究报道,PcSec62是否参与调控辣椒疫霉胁迫响应、生长及致病有待研究。本研究中,本研究团队对PcSec62进行基因功能分析,系统研究PcSec62在调节辣椒疫霉生长、发育、非生物胁迫响应和致病性方面的作用。
-
本研究所用的辣椒疫霉野生型菌株LT1534,大肠杆菌(Escherichia coli)菌株DH5α由本实验室保存;试验辣椒品种HUNCB226由海南大学汪志伟教授团队馈赠;基因编辑载体pYF515与pBlucscript Ⅱ KS+(pBS)由本实验室保存。
-
新活化至稳定生长的LT1534菌株转接至TA液体培养基[TA培养基:过滤番茄汁300 mL,加入纯水定容至3 L,并加入3.5 g 碳酸钙(CaCO3)],25 ℃ 120 r·min−1培养3 d后收集菌丝体;同时将V8平板(V8培养基:过滤V8果汁300 mL,加入纯水定容至3 L,并加入3.5 g CaCO3,每升加15 g琼脂粉)上培养3 d的LT1534进行光照诱导产孢3~5 d,收集孢子囊及游动孢子。取直径5 mm的菌丝块接种至5~6片真叶龄期的辣椒叶片上,置于黑暗条件侵染24 h,移去菌丝块按正常光周期培养;从接种开始计时,收集侵染12、24、48和72 h的叶片样本。利用RNA提取试剂盒提取菌丝体、孢子囊、游动孢子及侵染叶片样本总 RNA并测定浓度,取1 μg总 RNA利用逆转录试剂盒反转录为cDNA。用NCBI在线网站(
https://www.ncbi.nlm.nih.gov/tools/primer-blast/ )设计PcSec62特异性qPCR引物qPCR-F和qPCR-R(表1),以cDNA为模板Tublin B为内参, 用qPCR-F和qPCR-R进行qRT-PCR定量检测,每个样品试验重复3次,每组样品3个生物学重复,扩增结果用2−ΔΔCt法计算其相对表达量[20],并进行单因素方差显著性统计分析。名称
Primer序列(5′—3′)
Sequences (5′—3′)用途
AmplificationSec62-sgRNA1F CTAGCAGAGGACTGATGAGTCCGTGAGGACGAAACG
AGTAAGCTCGTCGTCCTCTATTCATACTGCAGApYF515载体 Sec62-sgRNA1R AAACTATGCAGTATGAATAGAGGAGACGAGCTTAC
TCGTTTCGTCCTCACGGACTCATCAGTGCAGAGpYF515载体 Sec62-sgRNA2F CTAGCTCCTCGCTGATGAGTCCGTGAGGACGAAAC
GAGTAAGCTCGTCCGAGGATGAAGTGGTAGACCpYF515载体 Sec62-sgRNA2R AAACGGTCTACCACTTCATCCTCGGACGAGCTTA
CTCGTTTCGTCCTCACGGACTCATCAGCGAGGAGpYF515载体 HPH-sgRNA1F CTAGCCGGAATCTGATGAGTCCGTGAGGACGAAACGAGTAA
GCTCGTCATTCCGGAAGTGCTTGACATpYF515载体 HPH-sgRNA1R AAACATGTCAAGCACTTCCGGAATGACGAGCTTACTCGTTT
CGTCCTCACGGACTCATCAGATTCCGGpYF515载体 HPH-sgRNA2F CTAGCTTGTCCCTGATGAGTCCGTGAGGACGAAACGAGT
AAGCTCGTCGGACAATGGCCGCATAACAGpYF515载体 HPH-sgRNA2R AAACCTGTTATGCGGCCATTGTCCGACGAGCTTACTCGT
TTCGTCCTCACGGACTCATCAGGGACAAGpYF515载体 AF TATCGATAAGCTTGATATCG
CACGAACAGCGAGGCTTTCpBS载体 AR GTGAGTTCAGGCTTTTTCAT
AGTGCTCCAAAAGGTTCTGpBS载体 BF CCGAGGGCAAAGGAATAG
ACGGCACCCTAGTAGGCAGpBS载体 BR CGGCCGCTCTAGAACTAGTG
ATGCTGACATGGGTGTGGpBS载体 HPHF ATGAAAAAGCCTGAACTC HPH片段扩增、PCR验证 HPHR CTATTCCTTTGCCCTCGG HPH片段扩增、PCR验证 ORF-F TCCGCAAGATGGCTGACT PcSec62片段扩增、PCR验证 ORF-R ACTCGTCAAAGTCCGTGGG PcSec62片段扩增、PCR验证 qPCR-F TGGGAGACGGAGAAGGGA qPCR 检测 qPCR-R CTAAACGGTCGCCACGGA qPCR 检测 Table 1. Primers used in the experiment
-
参照Wang等[21]和Qiu等[22]的方法,用EuPaGDT在线网站(
http://grna.ctegd.uga.edu/batch_tagging.html )及Primer 5.0设计试验所需引物(表1)。经体外合成Sec62-sgRNA1与Sec62-sgRNA2双链片段,利用T4 DNA连接酶将其连接到sgRNA表达载体pYF515;以LT1534基因组DNA为模板扩增PcSec62上下游1 000 bp同源臂,同时扩增潮霉素磷酸转移酶(Hygromycin phosphotransferase,HPH)基因片段;将扩增获得的片段连接到pBS载体上。制备LT1534原生质体,通过PEG介导法将重组pYF515及pBS质粒转入原生质体;过夜黑暗培养后加入含终浓度为100 μg·mL−1氨苄青霉素(ampicillin)、60 μg·mL−1遗传霉素(G418)的PAM培养基(PM/PAM培养基:取700 mL豌豆汁,加入91.1 g D-mannitol,1 g CaCl2,2 g CaCO3,用纯水定容至1 L,每升加15 g琼脂粉),待长出菌丝后加入含终质量浓度为100 μg·mL−1氨苄青霉素、100 μg·mL−1 G418和50 μg·mL−1潮霉素的V8培养基继续培养,至平板表面长出菌丝后挑取转化子进行PCR验证,并送至生工生物工程(上海)股份有限公司测序分析。参照上述基因敲除方法,合成HPH-sgRNA1与HPH-sgRNA2,连接到pYF515载体,同时以LT1534基因组DNA为模板扩增PcSec62上下游同源臂与PcSec62片段连接到pBS载体,获得重组载体。通过PEG介导法将重组pYF515和pBS载体转入辣椒疫霉ΔPcsec62的原生质体中,通过G418及氨苄抗性筛选回补转化子,并进行PCR验证及测序分析,获得回补菌株ΔPcsec62-C。
-
取直径5 mm辣椒疫霉野生型LT1534、pBS和pYF515空载体(empty vector,EV)转化菌株EV、突变体ΔPcsec62及回补菌株ΔPcsec62-C的菌丝块转接至V8培养基,25 ℃黑暗培养,每12 h测量1次菌落直径,72 h拍照记录,每个处理3个生物学重复。分别将供试菌株转接至TA液体培养基静置培养3 d,用光学显微镜10 ×和40 ×倍镜观察菌丝形态,统计100根菌丝每200 μm菌丝膨大体的数量,并进行单因素方差显著性统计分析。
-
将辣椒疫霉菌株LT1534、EV、ΔPcsec62、ΔPcsec62-C培养平板进行光照诱导产孢,3 d后收集孢子囊,制备悬浮液过滤并定容至1 mL。吸取10 μL孢子囊悬浮液至血球计数板上计数。每个供试菌株3个生物学重复,每管孢子囊悬浮液计数3次,通过单因素方差分析法进行显著性统计分析。
-
分别将盐胁迫剂氯化钠(NaCl)、钾离子胁迫剂氯化钾(KCl)、细胞膜胁迫剂十二烷基硫酸钠(SDS)、渗透胁迫剂山梨醇(Sorbitol)、内质网胁迫剂二硫苏糖醇(DTT)和细胞壁胁迫试剂刚果红(CR)添加至V8培养基中,分别调整终质量分数为0.2% NaCl、0.2% KCl、0.004% SDS、0.2 mol·L−1 Sorbitol、0.2 mol·L−1 DTT、200 μg·mL−1 CR[23-25]。将辣椒疫霉菌株LT1534、EV、ΔPcsec62、ΔPcsec62-C转接至含各胁迫剂的V8培养基上进行观察,72 h后测量菌落直径并拍照记录。每个处理做3个生物学重复,测量结果进行单因素方差分析法显著性统计分析。
-
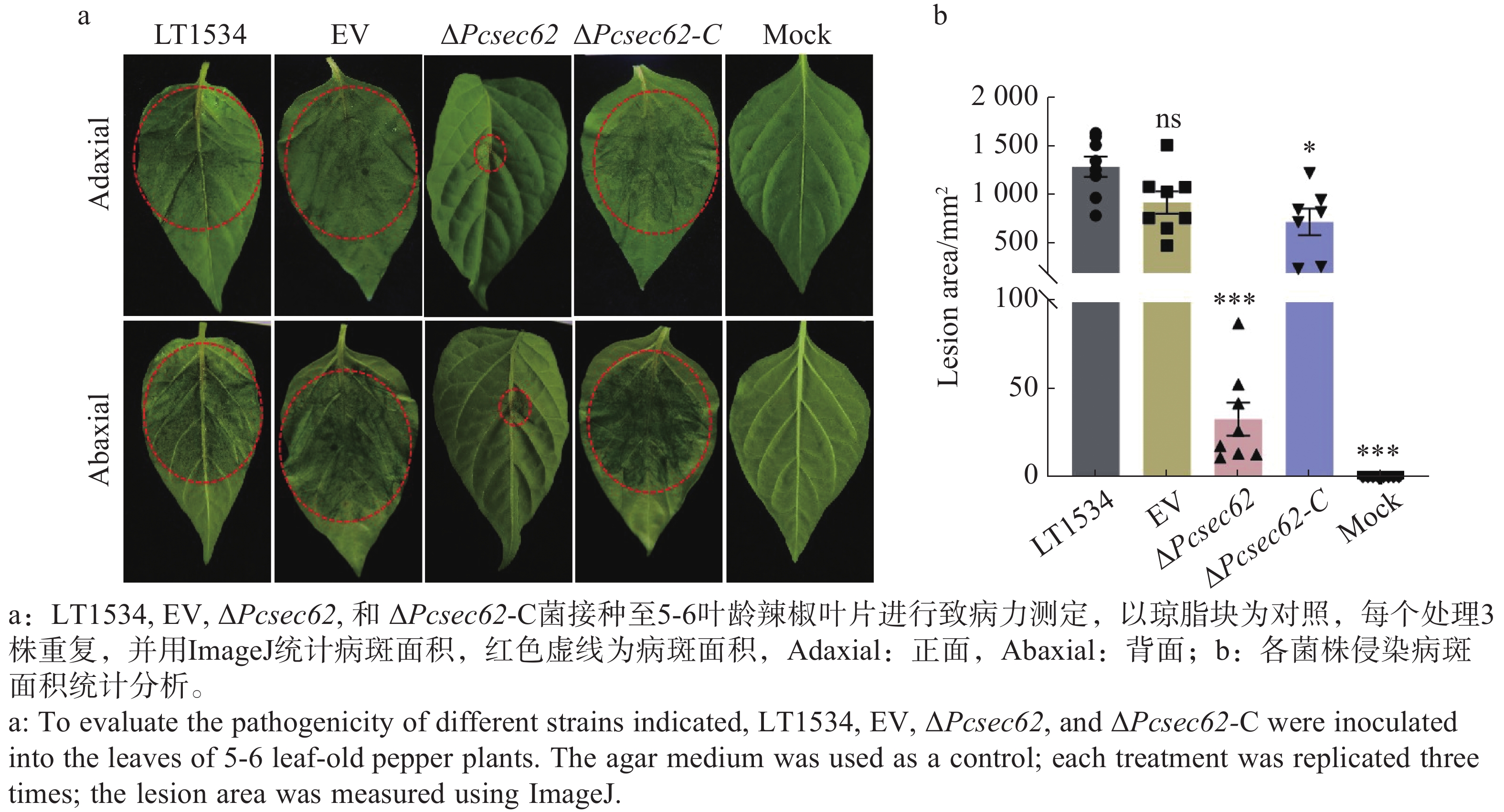
取直径为5 mm的LT1534、EV、ΔPcsec62、ΔPcsec62-C菌丝块接种至5~6片真叶龄期的辣椒叶片,以空白琼脂块为对照(Mock);黑暗培养24 h后移除菌块继续培养48 h观察致病情况,并用ImageJ软件统计病斑面积,每个处理3株重复,并通过单因素方差分析法进行显著性统计分析。
-
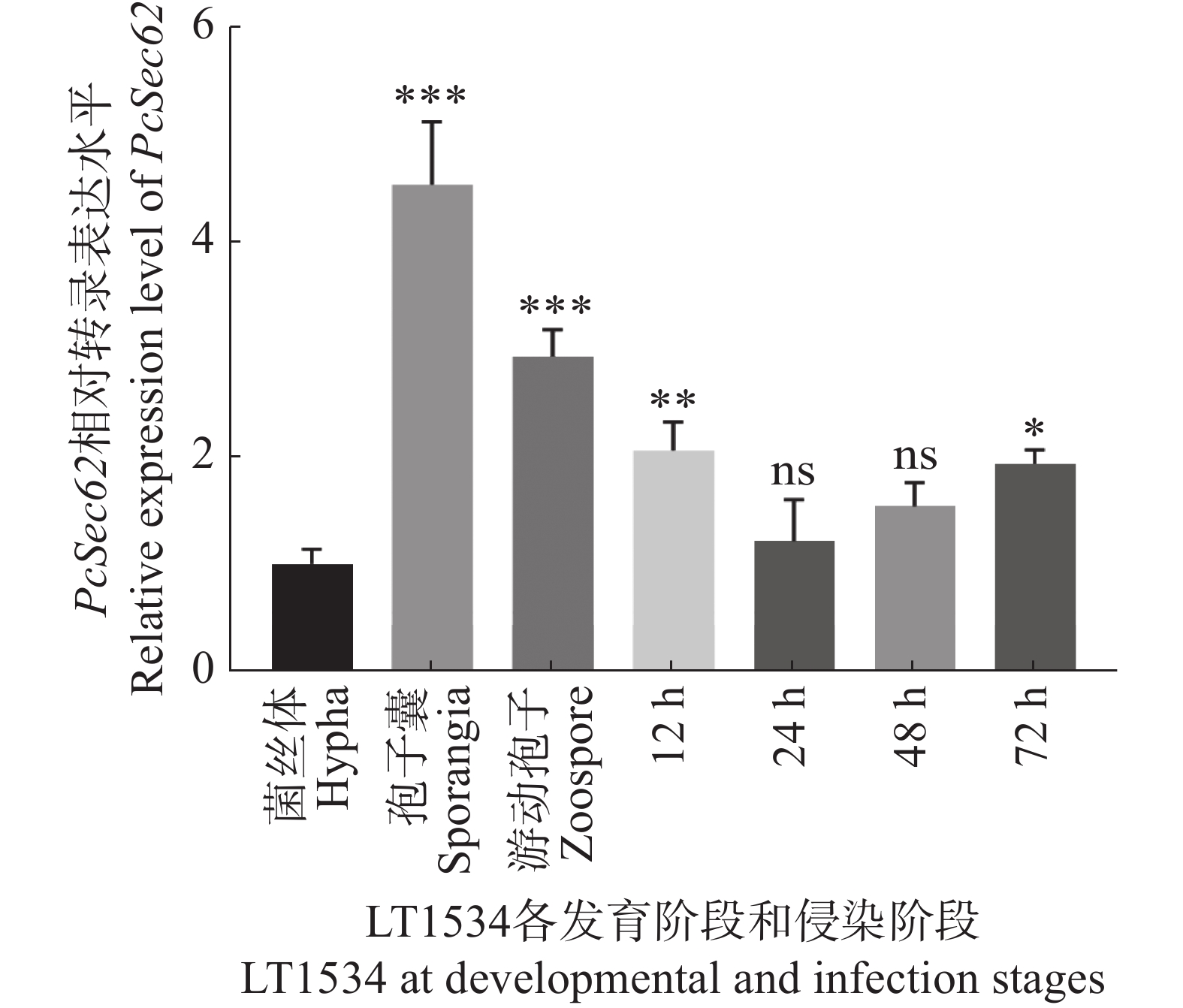
为确定PcSec62是否参与调控辣椒疫霉的生长发育及致病性,提取菌丝、孢子囊和游动孢子及辣椒疫霉侵染辣椒叶片4个时间点样本的总RNA进行qRT-PCR定量分析。结果表明,在已检测的辣椒疫霉3个发育阶段中,PcSec62在孢子囊中表达量最高,游动孢子次之,菌丝体的表达量最低;而在侵染阶段,与纯菌丝体相比,PcSec62在侵染12 和72 h的表达量显著上调(图1),该结果表明PcSec62可能参与调控辣椒疫霉的发育和致病阶段。
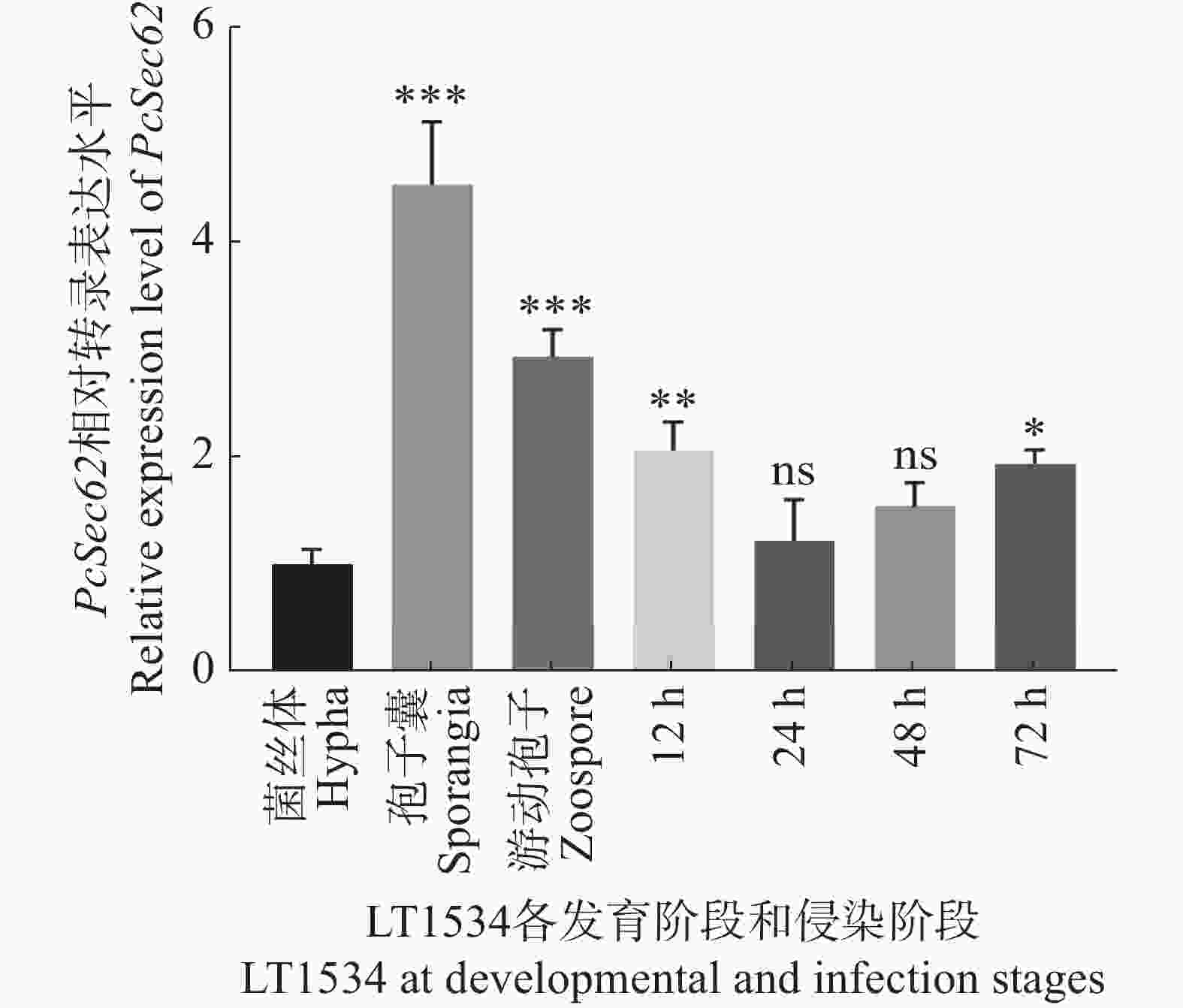
Figure 1. qRT-PCR detection of the relative mRNA transcript levels of PcSec62 during the development and infection stages of P. capsici.
-
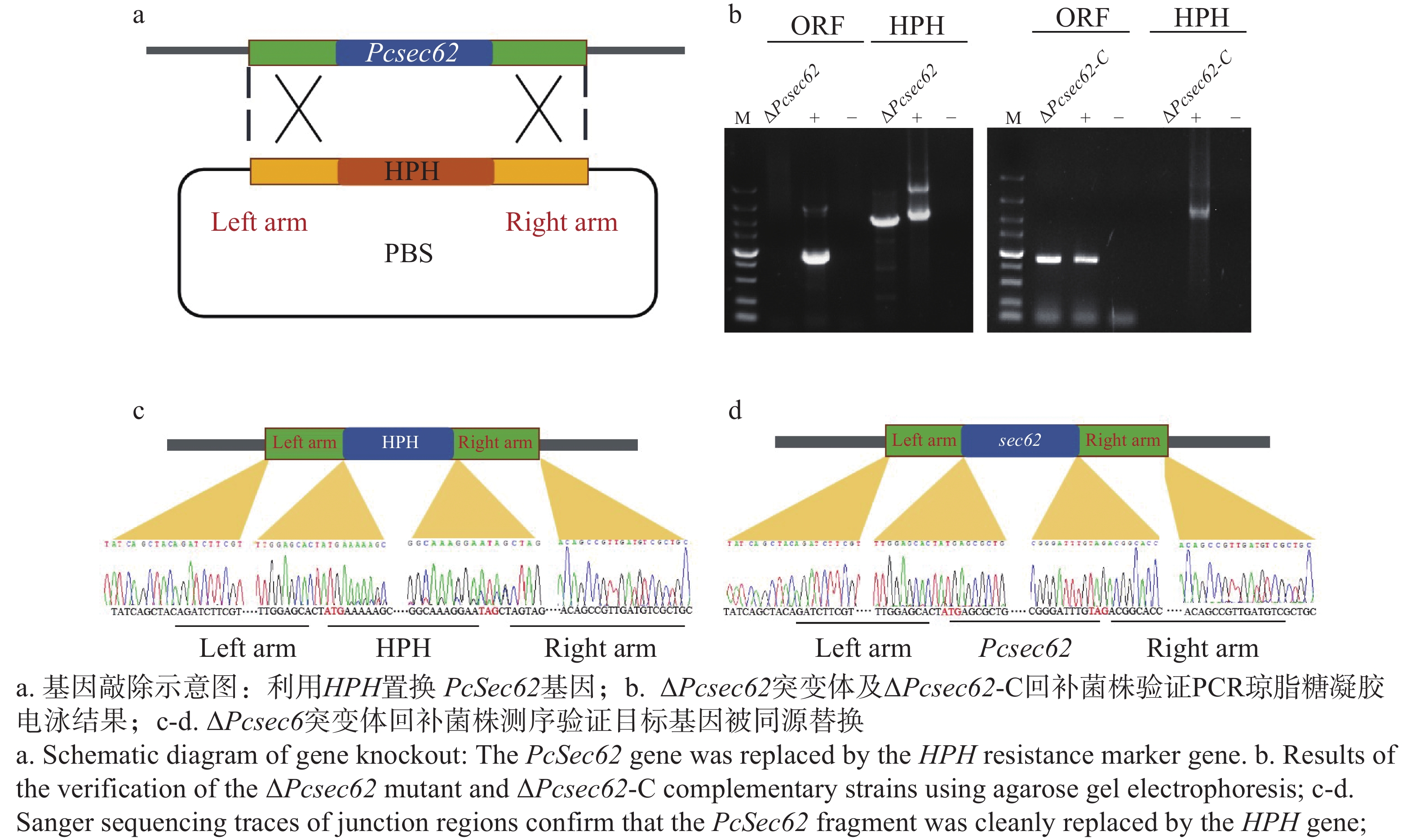
为研究PcSec62基因在辣椒疫霉中的功能,利用CRISPR/Cas9基因编辑系统,用HPH基因对PcSec62进行同源替换(图2-a),对获得的转化子进行PCR验证。结果显示,ΔPcsec62敲除转化子中PcSec62基因检测无条带,而HPH基因检测有条带,表明PcSec62基因被HPH基因成功置换,证明获得PcSec62候选完全敲除突变体(图2-b)。经测序分析证实PcSec62基因已成功被HPH基因替换,证明获得ΔPcsec62突变体(图2-c)。随后,对ΔPcsec62突变体进行PcSec62基因原位回补,ΔPcsec62-C回补转化子中HPH基因检测无条带,而PcSec62基因检测有条带。通过PCR检测和测序分析证实获得了原位回补菌株ΔPcsec62-C(图2-b,d)。
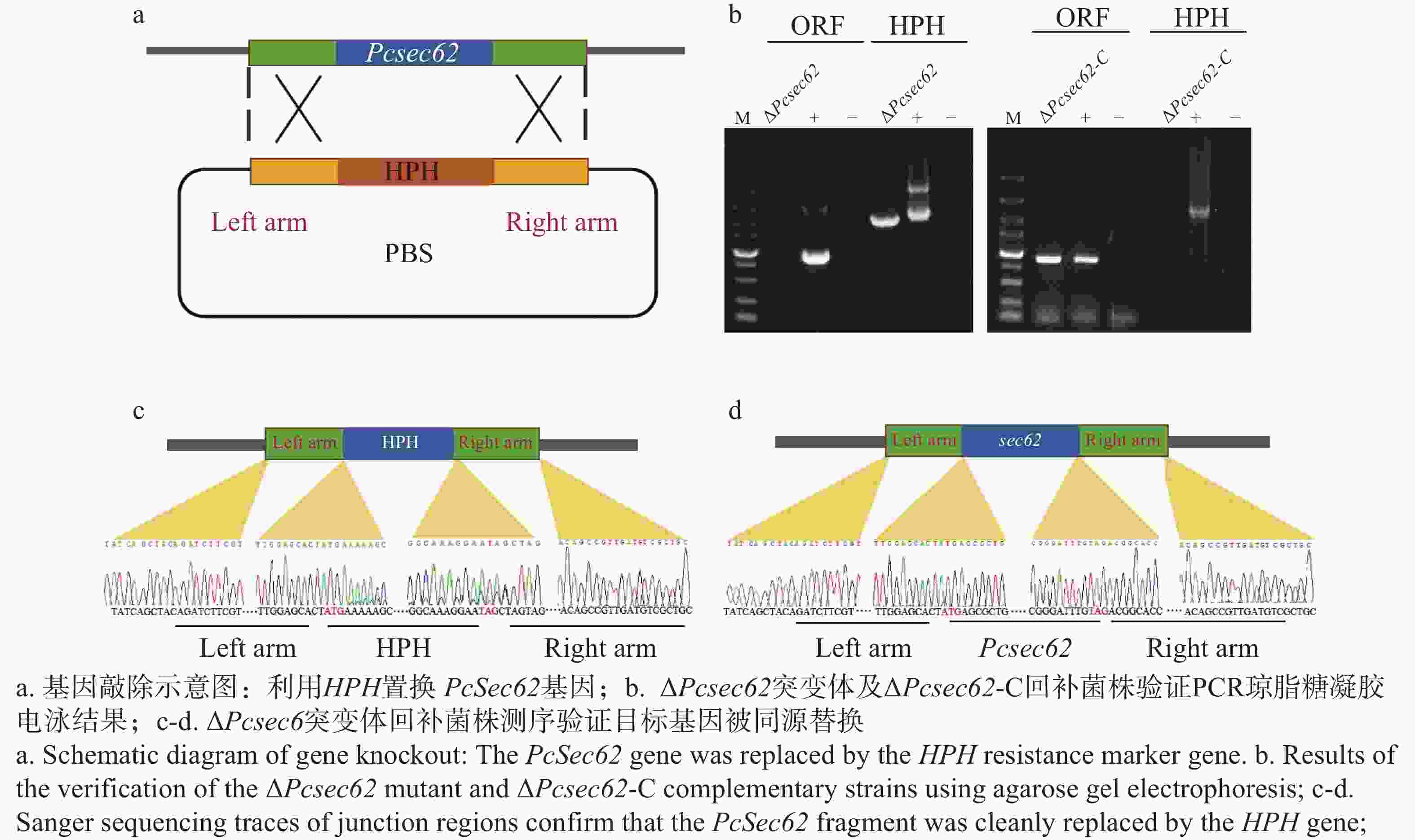
Figure 2. Verification of PcSec62 gene knockout and complementation in P. capsici strains.
-
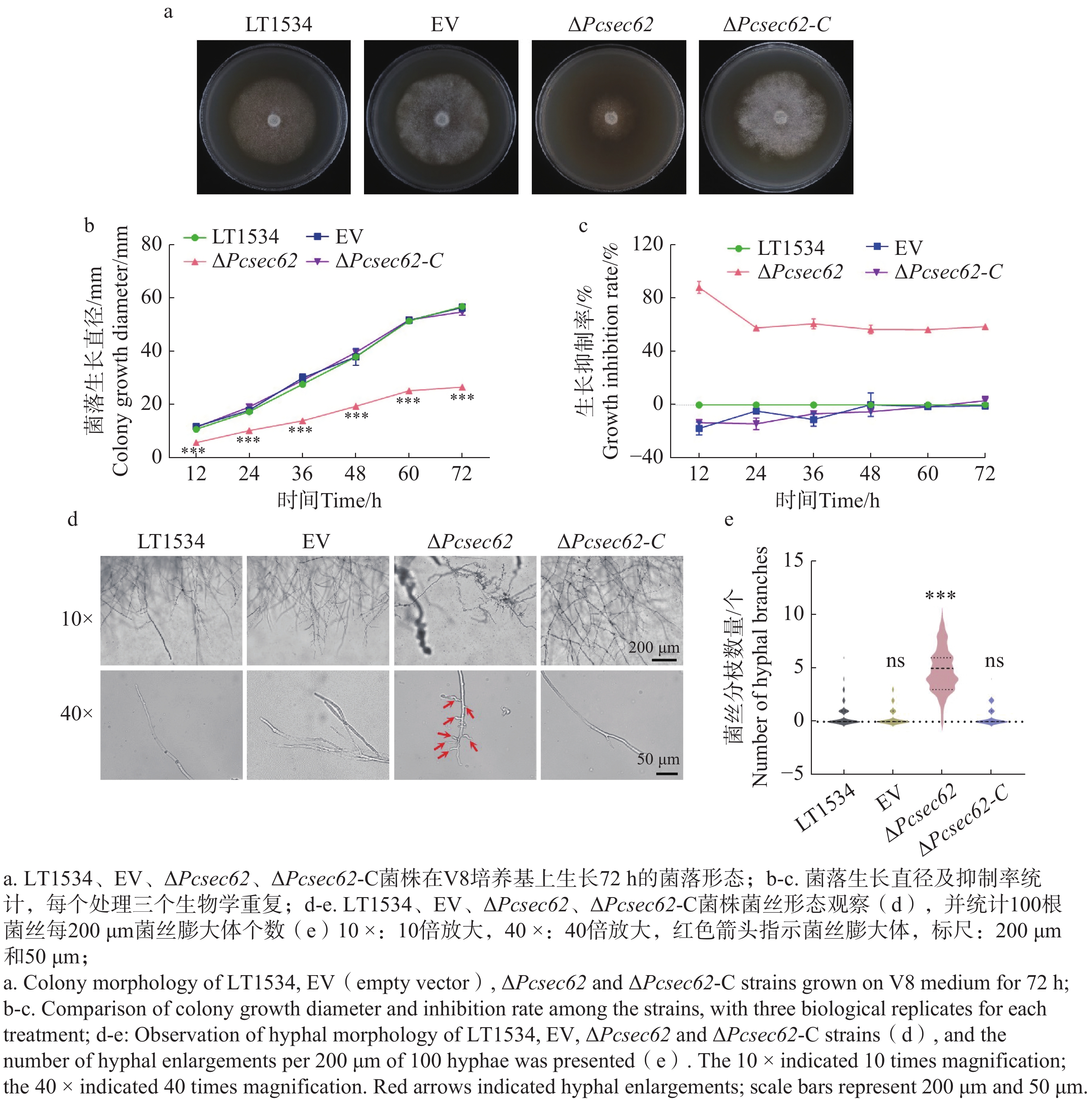
为明确PcSec62是否影响辣椒疫霉生长和形态变化,在V8培养基上分别接种LT1534、EV、ΔPcsec62和ΔPcsec62-C菌株;每12 h测量1次直径,72 h后拍照记录并统计分析。结果表明,EV和LT1534菌丝生长速率基本相同;与LT1534相比,ΔPcsec62生长速度显著降低,ΔPcsec62-C菌株生长速率恢复到LT1534的水平(图3-a,b,c),表明PcSec62参与调控辣椒疫霉的菌丝生长。进一步对供试菌株菌丝尖端的形态进行观察。结果表明:ΔPcsec62菌丝尖端的菌丝膨大体异常增加(图3-c),通过统计100根菌丝体每200 μm的菌丝膨大体的数量,发现ΔPcsec62突变菌丝尖端膨大体的数量显著高于LT1534、EV。回补PcSec62基因后,菌丝膨大体数量恢复至野生型水平(图3-d)。表明PcSec62可能通过抑制辣椒疫霉菌丝极性生长,从而影响菌丝生长速率。
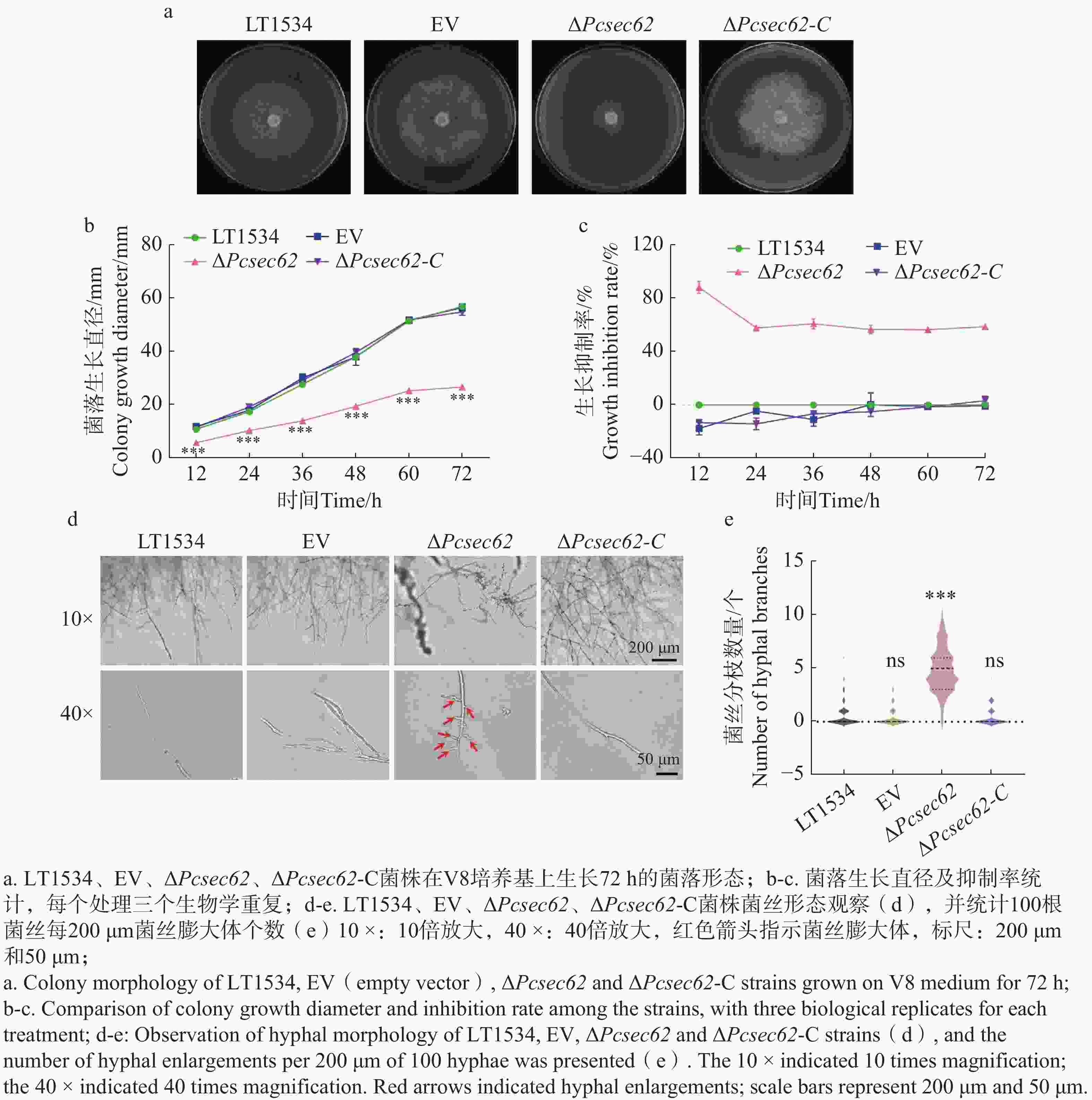
Figure 3. Effect of ΔPcsec62 on colony and hyphal morphology of P. capsici
-
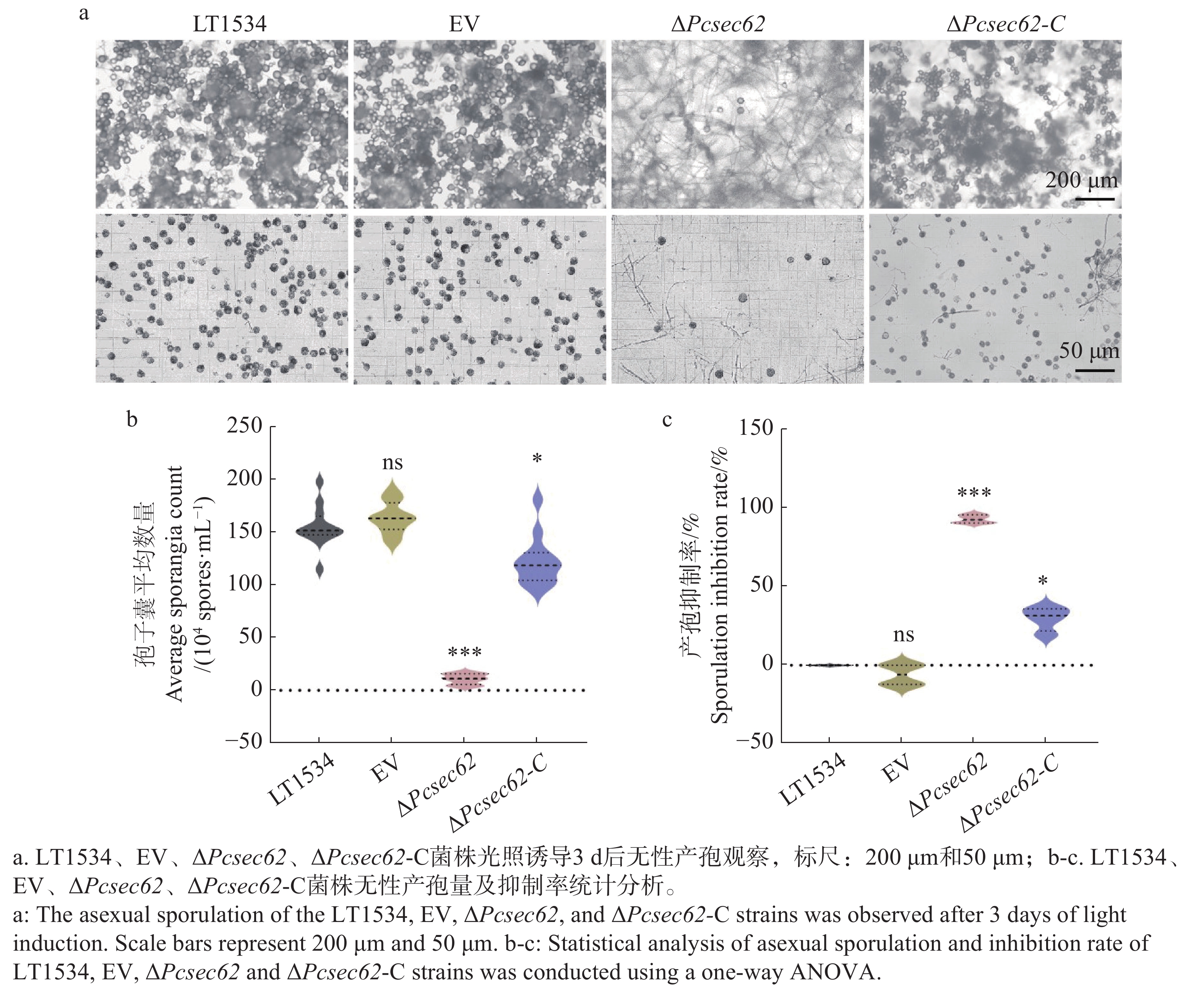
为研究PcSec62对辣椒疫霉产孢能力的影响,对LT1534、EV、ΔPcsec62、ΔPcsec62-C菌株进行光照诱导并对孢子囊的数量进行统计。结果显示:LT1534、EV、ΔPcsec62、ΔPcsec62-C菌株孢子囊数量分别为1.552×106、1.638×106、1.075×105、1.225×106个·mL−1,相比于LT1534和EV菌株,ΔPcsec62突变体孢子囊数量显著下降,而回补菌株ΔPcsec62-C孢子囊产量恢复到约野生型水平的80%(图4-a,b)。结果表明PcSec62参与调控辣椒疫霉的无性繁殖。
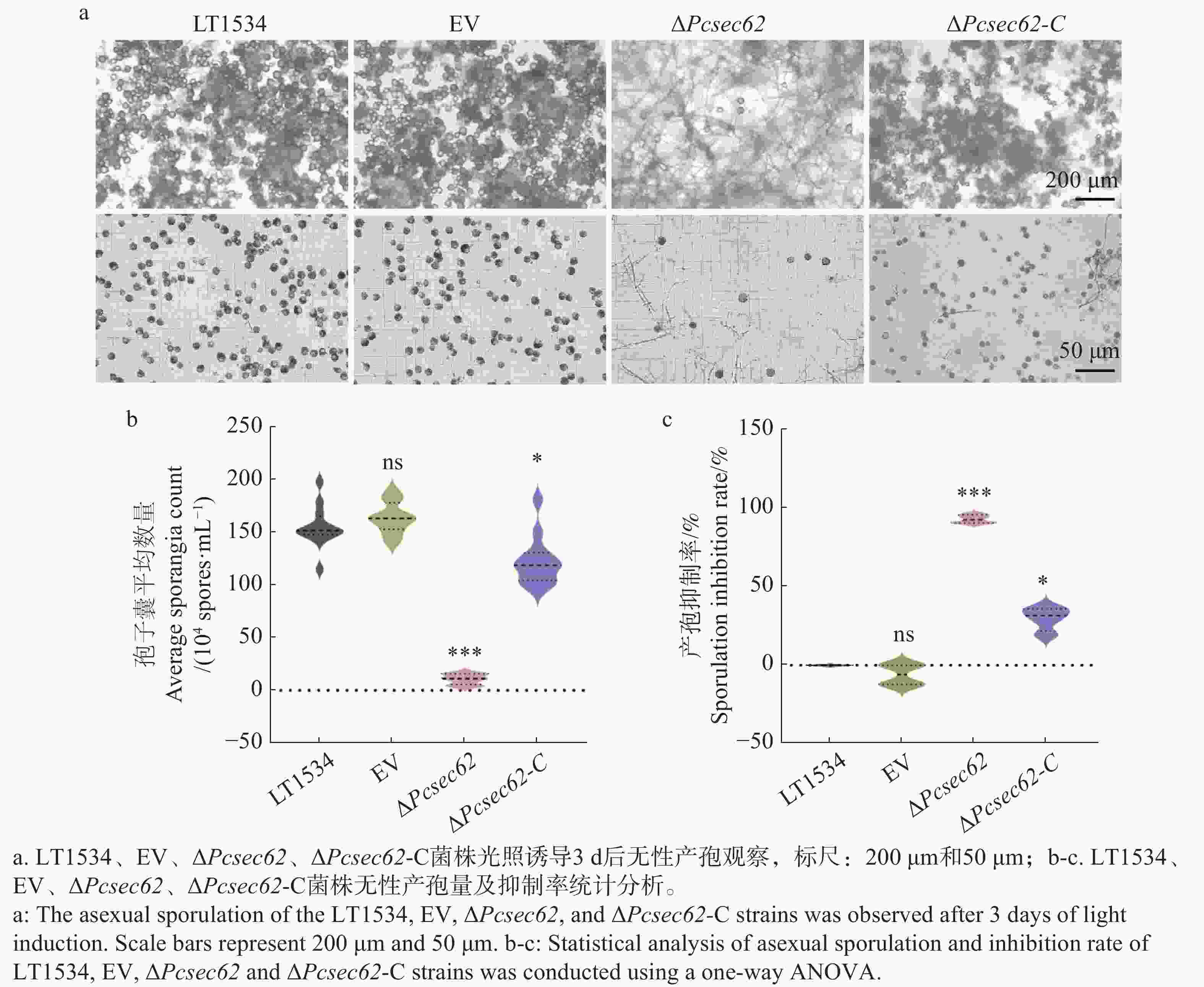
Figure 4. Impact of ΔPcsec62 on asexual sporulation in P. capsici
-
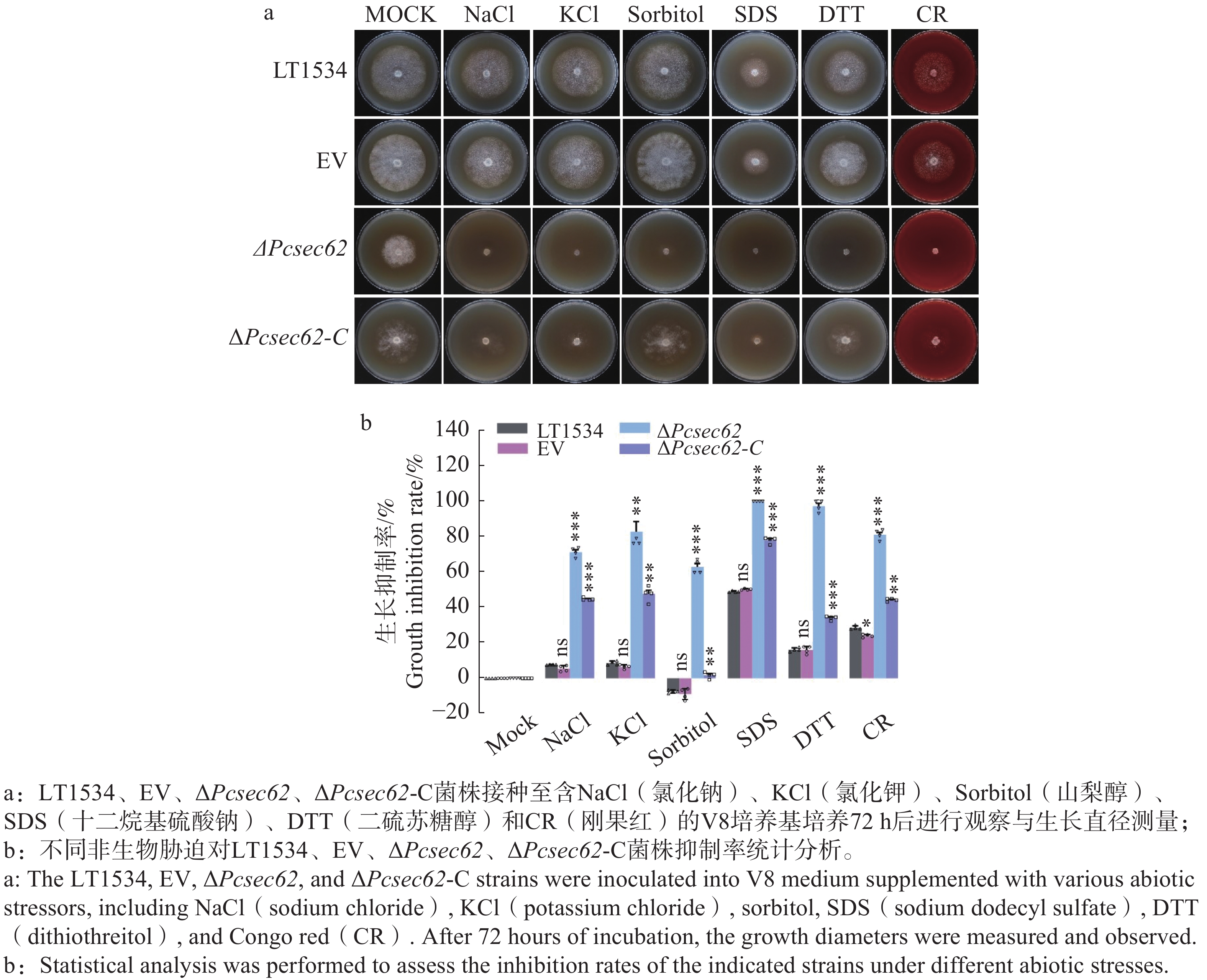
为明确PcSec62基因在非生物胁迫反应中的作用,将LT1534、EV、ΔPcsec62、ΔPcsec62-C菌株分别接种至含NaCl、 KCl、SDS、Sorbitol、DTT、CR胁迫剂的培养基进行观察,72 h后测量直径、拍摄照片并计算分析其抑制率(图5-a)。结果表明,与LT1534相比,ΔPcsec62对各胁迫试剂的耐受性显著降低,对ΔPcsec62的抑制率NaCl为71.29%,KCl为82.90%,Sorbitol为62.97%,SDS为100.00%,DTT为97.33%,CR为81.12%;回补PcSec62基因后ΔPcsec62-C对非生物胁迫耐受能力部分得到恢复(图5-b),表明PcSec62可能参与辣椒疫霉对非生物胁迫的调控。
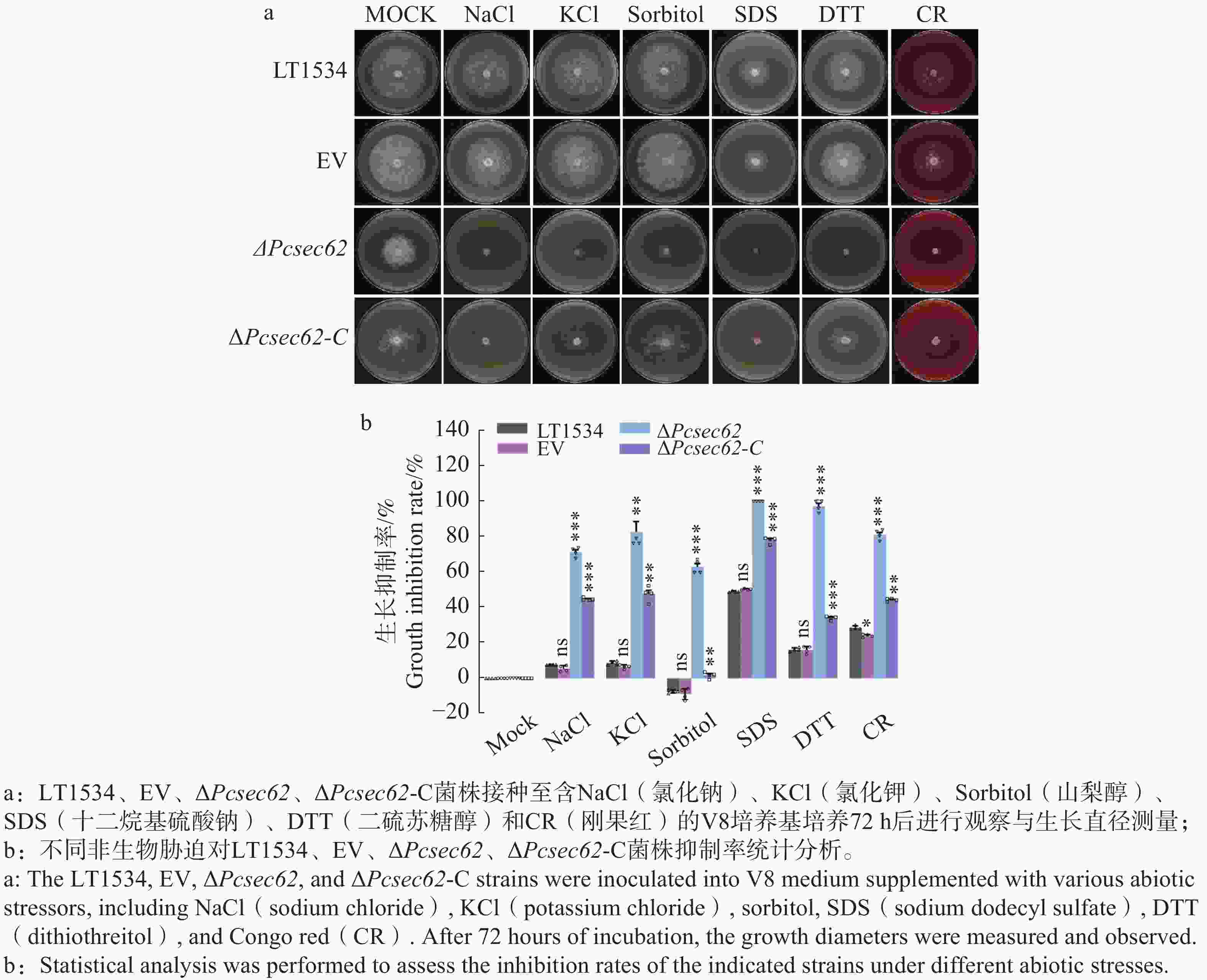
Figure 5. PcSec62 is involved in the regulation of abiotic stress tolerance of P.capsici
-
为了确定PcSec62对辣椒疫霉致病性的影响,以琼脂块作为空白对照,分别将LT1534、EV、ΔPcsec62、ΔPcsec62-C接种到辣椒叶片表面进行侵染(图6-a)。结果表明与LT1534和EV对照相比,ΔPcsec62侵染叶片后造成的病斑面积显著变小,而回补PcSec62基因后ΔPcsec62-C 致病能力基本恢复至LT1534水平(图6-b)。这表明PcSec62在调节辣椒疫霉的侵染及致病过程中起着重要作用。
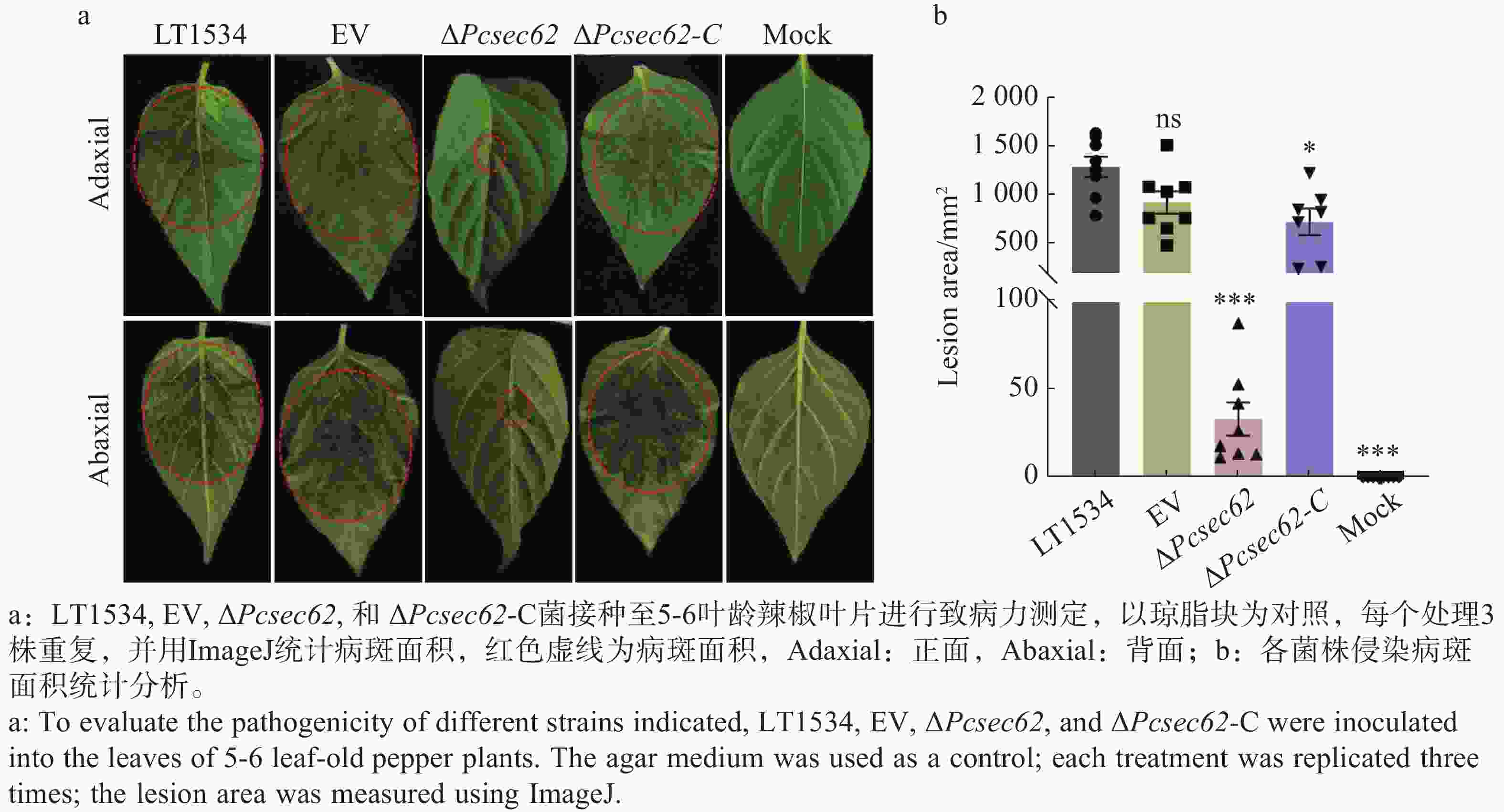
Figure 6. Effect of PcSec62 on the pathogenicity of P. capsici
-
Sec62是内质网膜易位子复合体的重要成员,定位于内质网膜,参与调节内质网应激恢复,介导内质网中未折叠蛋白以及错误折叠蛋白向溶酶体的运输维持细胞稳态等多种生命过程[26]。在本研究中,辣椒疫霉PcSec62在各生长发育及侵染阶段中菌体与纯菌丝体相比在转录水平上差异显著,在孢子囊及致病过程中显著上调表达,这表明PcSec62对辣椒疫霉生长发育及致病性至关重要。在植物中,AtSec62缺失导致拟南芥生长受损,花粉异常和育性下降[17-18];辣椒疫霉PcSec62缺失后菌丝生长受到抑制,产孢能力几乎丧失,而回补菌株的菌丝生长和产孢表型基本恢复至野生型水平,这证明辣椒疫霉PcSec62与AtSec62功能类似,对生长及发育至关重要。这可能是由于Sec62作为易位蛋白复合体组分,在促进分泌蛋白与膜蛋白的易位和分泌所引起[27]。有意思的是,PcSec62缺失后菌丝生长受阻,菌丝膨大体数量显著增加。在丝状真菌中,菌丝生长是通过菌丝尖端凝集新的质膜,伴随着分泌囊泡的胞吐作用合成新的细胞壁等组分引起[28-29]。这可能是PcSec62的缺失破坏了分泌途径,间接影响菌丝体尖端的生长,从而抑制菌丝的营养生长。内质网相关基因FgErv14和MoDer1参与真菌的无性繁殖[30-31],RTN1则可调控有性生殖过程,表明内质网相关因子对生殖发育至关重要[32]。本研究中,PcSec62缺失导致辣椒疫霉几乎完全丧失产孢能力,这可能与PcSec62缺失破坏易位复合体,影响生长发育相关蛋白加工所致。本研究ΔPcsec62-C回补菌株在孢子囊产量及非生物胁迫表型未完全恢复至野生型水平。这可能是辣椒疫霉为二倍体而ΔPcsec62-C回补未获得纯合互补菌株导致的干扰现象。
Sec62是重要的内质网应激调节因子,参与内质网的应激恢复[16]。本研究发现,PcSec62的缺失使辣椒疫霉对钠盐、钾离子、细胞壁、细胞膜等非生物胁迫更敏感,但回补后敏感性得到部分缓解。尽管本研究结果尚未阐明PcSec62对非生物胁迫的调控机制,但可以证实PcSec62在辣椒疫霉对环境胁迫中起着重要作用。此外,PcSec62的缺失导致辣椒疫霉的致病性几乎完全丧失,这可能是PcSec62直接影响辣椒疫霉生长及产孢等表型,也可能Sec62的缺失破坏内质网易位系统及分泌系统,从而影响大量致病相关效应蛋白合成或分泌受阻,间接导致辣椒疫霉致病力下降[33-35]。
-
本研究通过对PcSec62在辣椒疫霉生长发育及侵染阶段的转录表达及生长、产孢和致病性生物学功能分析,证实PcSec62参与调控辣椒疫霉的生长发育、胁迫应答及致病性。本研究结果对阐明内质网途径介导的致病机制具有重要意义,但具体机制仍有待进一步研究。
PcSec62 is involved in regulating the growth, development and virulence of Phytophthora capsici
DOI: 10.15886/j.cnki.rdswxb.20240170
- Received Date: 2024-11-10
- Accepted Date: 2025-01-10
- Rev Recd Date: 2024-12-05
- Available Online: 2025-03-11
- Publish Date: 2026-03-25
-
Key words:
- Phytophthora capsici /
- PcSec62 /
- endoplasmic reticulum stress /
- growth and development /
- virulence
Abstract: The endoplasmic reticulum (ER) serves as a crucial site for protein processing in eukaryotic cells. Sec62, an essential component of the ER translocation complex, plays a significant role in growth, development, and stress regulation. An attempt was made to examine the transcription levels of the PcSec62 gene at various growth stages and during pathogenic processes. The PcSec62 gene was knocked out by using CRISPR/Cas9 gene editing to produce knockout mutants (ΔPcsec62) and a complementary strain (ΔPcsec62-C). The results showed that the transcriptional expression of PcSec62 significantly increased during the sporangia and infection stages. The ΔPcsec62 mutants exhibited notably reduced growth and sporulation abilities, alongside stunted hyphal growth. Additionally, the ΔPcsec62 strain showed significantly low tolerance to abiotic stress and reduced pathogenicity. These findings indicate that PcSec62 is involved in regulating the growth, development, abiotic stress responses, and pathogenicity of Phytophthora capsici.
| Citation: | Huang Yuyuan, Ye Qianqian, Chen Qinghe, Liang Qifu. PcSec62 is involved in regulating the growth, development and virulence of Phytophthora capsici[J]. Journal of Tropical Biology, 2026, 17(2): 314-323. doi: 10.15886/j.cnki.rdswxb.20240170

|
 DownLoad:
DownLoad: