
-
橘小实蝇[Bactrocera dorsalis (Hendel)]又名东方果实蝇、柑橘小实蝇,隶属于双翅目(Diptera)实蝇科(Tephritidae)果实蝇属(Bactrocera),是一种危害严重的世界性检疫害虫[1-2]。橘小实蝇起源于热带和亚热带地区,1912年在中国台湾岛被首次记录[3],其具有寄主范围广、成虫飞行能力强、气候耐受性高等特点,现已广泛分布于中国和东南亚、印度次大陆、夏威夷群岛等区域[4]。橘小实蝇危害性主要来源于幼虫的取食,橘小实蝇雌虫会将卵产于寄主植物果皮下,幼虫孵化后大量取食果肉,造成果实腐烂、脱落,严重影响果实产量和质量[5-6],同时严格的检疫政策也限制了橘小实蝇疫区果蔬产品的出口[7]。这些因素对农业生产造成了严重的经济损失[8-9]。
以引诱剂为基础的行为调控技术被认为是一种绿色的害虫防控策略,其主要是指以性信息素、寄主植物挥发物或食物源挥发性气味等为基础合成的人工饵剂,可用于诱集目标害虫,检测、调控害虫种群[10-12]。在橘小实蝇中,以雄性引诱剂甲基丁香酚(methyl eugenol,ME)为代表的雄性歼灭策略(male annihilation technique,MAT)和双性蛋白食诱剂为核心的蛋白诱饵策略(protein bait application technique,BAT)已在种群防控中发挥了重要的作用[13]。但实际应用上也存在一些不足,例如橘小实蝇雄虫能够多次交配,这导致了部分未诱捕雄虫仍能维持较高的雌性繁殖群体[14];此外,蛋白饵剂虽对雌雄虫均具有引诱作用,但在田间具有易降解、有效时间短、易受pH影响等缺点[15-17]。这些因素限制了MAT和BAT在田间实际的种群控制效果,因此有必要去开发更多高效、稳定的行为调控策略和产品。
昆虫嗅觉系统是引诱剂发挥功能的基础,是昆虫长期进化形成的高度敏锐的外周环境感受系统,它参与了昆虫觅食、求偶、交配、产卵等各个行为反应[18-19]。解析昆虫的嗅觉识别系统,将为害虫行为调控剂的改良和开发提供重要的理论基础。在昆虫中,触角是主要的嗅觉识别器官,气味分子首先要通过触角感器皮孔进入淋巴液,经气味结合蛋白运输至嗅觉受体神经元,再由神经元上的嗅觉受体将化学信号转化为电信号,传递至上游的触角嗅觉中枢触角叶,各种嗅觉信号在此处被初步加工后进一步通过投射神经元传递至嗅觉高级中枢,从而指导相应的行为反应[20-22]。因此,建立化学物质→嗅觉受体神经元→嗅觉中枢→行为的对应关系,是了解昆虫嗅觉感受系统的重点。橘小实蝇中虽已报道了大量的嗅觉相关蛋白,但这些分子靶标与神经和行为的对应关系仍不明确[23-25],且昆虫触角的感器数量往往众多,类型复杂,通过直接筛选来定位感器、嗅觉受体神经元与化合物的对应关系,往往耗时耗力;此外,传统的神经元染色标记技术在非模式昆虫的应用上也存在着挑战。为克服传统方法的局限性,科学家提出了基于泛神经元表达基因的转基因神经元标记技术[26]。泛神经元表达基因是指一类在神经系统中广泛表达的功能基因,它们几乎存在于所有神经元中,是神经元功能的基础[26-27]。科学家通过这类基因的转录调控区域在神经元上驱动特异性报告基因的表达,实现了化感神经元图谱的构建及对重要化合物感受神经元的标记和活性监测[26]。在模式昆虫黑腹果蝇(Drosophila melanogaster)、埃及伊蚊(Aedes aegypti)、嗜水按蚊(Anopheles coluzzii)中,通过这种技术已完成了对重要化合物感受图谱的绘制[26,28-30]。在这些昆虫中目前常用的目标基因主要包括Brp蛋白基因(bruchpilot,Brp)、神经元突触小泡结合蛋白基因(neuronal Synaptobrevin,nSyb)、突触结合蛋白1基因(synaptotagmin1,Syt1)和胚胎致死异常视蛋白基因(embryonic lethal abnormal vision,Elav)。其中,Brp是一种细胞骨架蛋白,主要存在于突触活性区[28];nSyb编码神经元突触小泡蛋白,参与神经传递过程的囊泡运输和融合[31-32];Syt1编码的突触结合蛋白-1是一种Ca2+感受器,可感知钙离子并介导突触小泡的释放,确保信号在神经元之间准确且高效地传递[33];Elav是一类神经元特异性RNA结合蛋白,对于神经元分化、成熟和神经系统的维持至关重要[34-35]。
橘小实蝇触角感器丰富,个体微小,开发高效的泛神经元表达工具将有助于橘小实蝇嗅觉系统的研究,但目前在此方向上还没有相关的基础。因此,本研究以果蝇中泛神经元研究常用的4种基因Brp、Syt1、nSyb、elav为基础,克隆了在橘小实蝇中的四类同源基因,并对它们的进化关系、基因组结构和外周表达模式进行了分析,以期为后续橘小实蝇泛神经元表达体系的构建提供基础。
-
本实验使用的橘小实蝇野生型品系由中国农业科学院深圳农业基因组研究所提供。饲养条件为温度(26±1) ℃,相对湿度(60±10)%,光周期光:暗=14 h:10 h,采用人工饲料饲养。幼虫饲料由新鲜香蕉、玉米粉、白砂糖、酵母及纤维素等组成。将幼虫养殖至老熟后,将其置于湿润的沙土中化蛹。化蛹4~5 d后晾干沙土,采用特制沙网将蛹筛出并置于亚克力养虫笼(18 cm × 24 cm × 14 cm)中羽化。羽化后前3 d将雌雄分开,饲养于实验专用的小型养虫笼(18 cm × 12.5 cm × 14 cm)中。成虫饲料为质量比1:1的白砂糖和啤酒酵母的混合物,饲养期间保证充足的水源。
选取12日龄左右的野生型成虫作为采集虫源。为保证顺利克隆出目标基因,本研究团队分别采集了4种神经元广泛分布的组织作为候选克隆模板,分别为脑、口器、下颚须、触角。脑、口器、下颚须、触角等组织的收集量分别为10对雌雄成虫的脑,100对雌雄成虫的下颚须、口器及触角。组织采集后置于无酶锆珠破碎管中,迅速放入液氮中速冻。−80 ℃冰箱中保存备用。
-
使用TRIzol试剂(Invitrogen,Carlsbad,CA,USA)提取各组织RNA,通过琼脂糖凝胶电泳和纳米微滴1000分光光度计(Thermo Scientific,Wilmington,DE)评估它们的完整性和浓度。以约1 µg RNA为模板,使用HiScript® Ⅲ 1st Strand c DNA Synthesis (+gDNA wiper) Kit(诺唯赞,南京,中国)进行第一链cDNA合成。所得cDNA稀释后于−20 ℃冰箱保存,以备后续PCR使用。
-
根据本实验室已有的橘小实蝇外周转录本数据(国家生物信息中心 GSA 数据库,PRJCA020830),利用TransDecoder v5.5.0鉴定了候选编码区,并采用Orthofinder v2.5.4[36]将从NCBI下载的黑腹果蝇(D. melanogaster)的全基因组蛋白序列与橘小实蝇的转录蛋白序列进行同源比对。根据比对结果,挑选与黑腹果蝇的基因DmBrp、DmnSyb、DmSyt1、Dmelav同源的候选BdornSyb、BdorSyt1、Bdorelav、BdorBrp序列。随后根据序列信息设计特异性引物,以合成的cDNA为模板PCR克隆4个基因。PCR产物经过琼脂糖凝胶电泳后,将目的基因条带切下,采用胶回收试剂盒(全式金,北京,中国)进行产物纯化。将纯化产物连接至Blunt载体(全式金,北京,中国),转化入Trans-T1感受态细胞中(全式金,北京,中国),37 ℃过夜培养。次日上午挑取白色单克隆菌落,进行菌液PCR验证插入片段。将阳性菌株进行扩繁并送至生工生物(上海,北京)测序。
-
使用ExPASy (
https://www.expasy.org/resources/protparam )平台上的ProtParam和ProtScale工具,分别计算了蛋白质的理论等电点、分子质量、不稳定指数、疏水指数以及平均疏水性指数。通过NCBI收集双翅目中已有报道相关基因的同源蛋白序列,以及部分外群序列,与上述所获得的橘小实蝇氨基酸序列一同使用 MAFFT v7.515[37]进行多序列比对,并使用 IQ-TREE v2.2.0.3[38]软件的 ModelFinder 选项选择最佳的核苷酸替代模型,以最大似然法构建系统发育树。采用MEME Suite (https://meme-suite.org/meme/index.html )分析各个基因同源序列的保守基序(motif)。 -
根据本实验室已有的外周神经组织的转录组数据,利用Blat[39]将橘小实蝇的基因BdornSyb、BdorSyt1、Bdorelav和BdorBrp比对至橘小实蝇参考基因组(国家生物信息中心 GSA 数据库,PRJCA020830),获得它们在基因组上的位置信息,为获得完整的基因结构信息,使用Samtools[40]截取每个基因比对位置上下游10K区域,再利用Hisat2 v2.2.1[41]将橘小实蝇外周神经组织的转录组数据比对至截取的基因组区域上,并将比对至目标基因组区域上的转录本reads通过Stringtie[42]进行组装,获得候选的转录本结构。使用Transdecoder (
https://github.com/TransDecoder/TransDecoder )预测转录本ORF,将比对到的基因组区域、转录组证据、转录本reads覆盖度以及预测的ORF导入IGV[43],进行IGV可视化,根据候选基因的基因组位置信息和转录组证据综合判断可信的基因结构。 -
根据已有组装的外周转录本数据,包括触角、口器、下颚须、前足、中足、后足、外生殖器共7种组织。采用Hisat2 v2.2.1[41]将转录本数据比对到实验室已有橘小实蝇参考基因组(NCBI项目号:PRJCA020830)。并采用featureCounts v2.0.1[44]计算原始表达量及TPM值,整合获得原始表达矩阵及TPM值矩阵。根据获得的橘小实蝇外周组织TPM表达矩阵,在其中提取目标基因BdornSyb、BdorSyt1、Bdorelav、BdorBrp的TPM值。
-
所有统计分析均使用Graph Pad Prism v8.0.1进行,所有重复数据结果均以平均值±标准误差表示。数据差异分析采用非配对t检验。
-
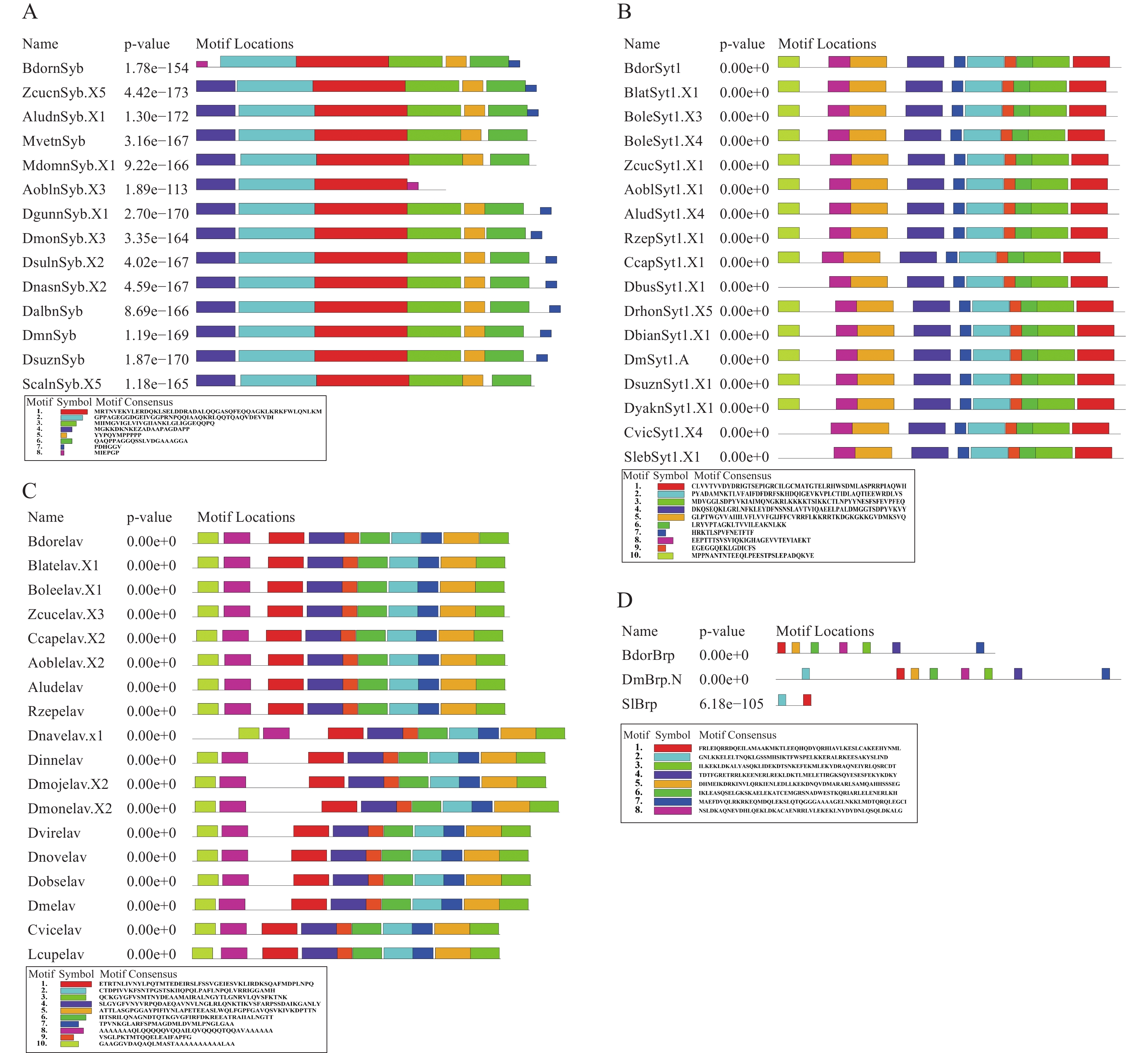
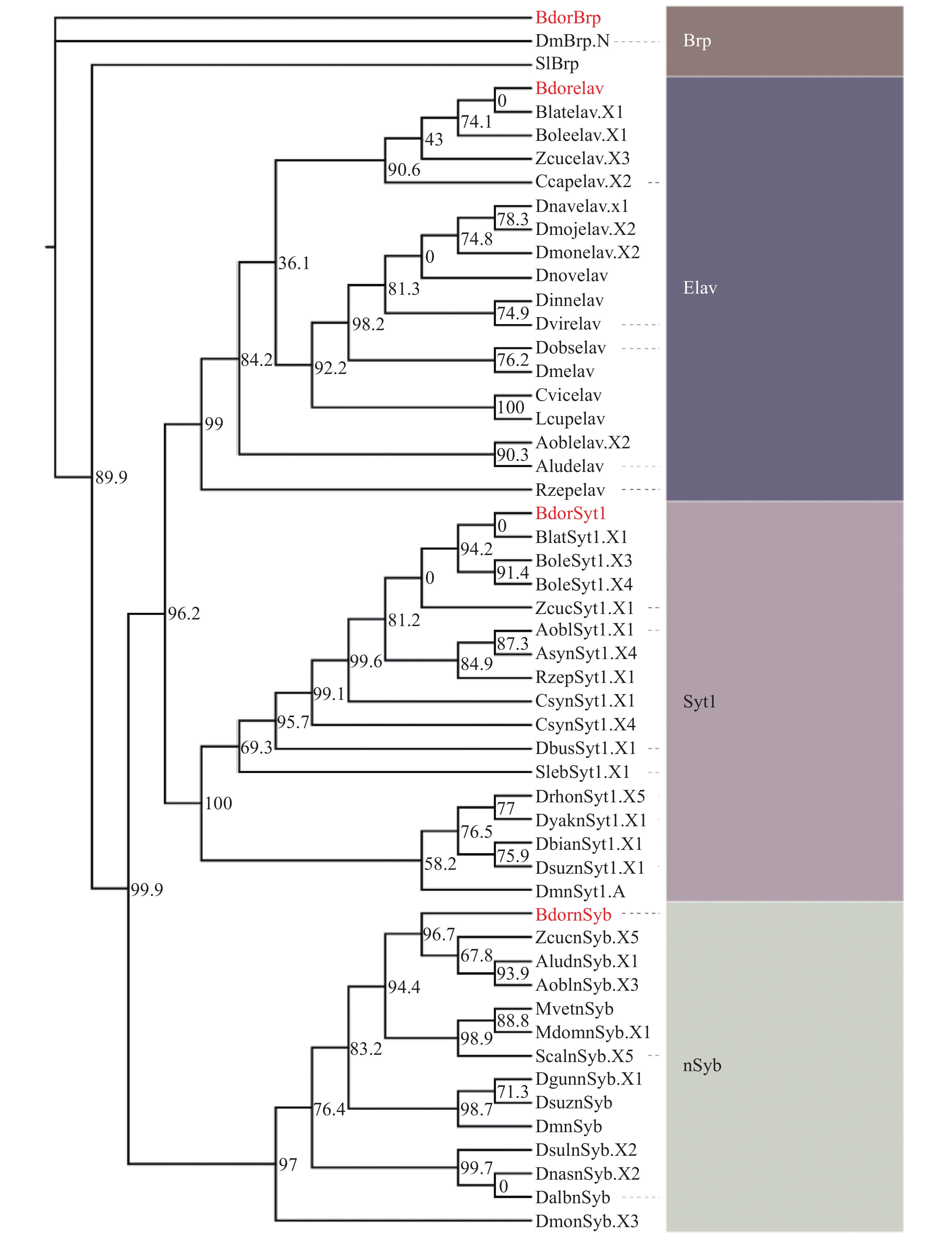
利用黑腹果蝇的4种基因DmBrp、DmnSyb、DmSyt1和Dmelav的全基因组蛋白序列,在实验室前期测序组装的橘小实蝇全基因组蛋白序列中均获得了高度同源序列(表1),并分别命名为BdorBrp、BdornSyb、BdorSyt1和Bdorelav。为进一步提高此类基因功能预测的准确性,选取此类基因在双翅目及其他有报道的相关物种中的序列,通过最大似然法(maximum likelihood method)构建了系统发育树,结果表明,13种nSyb蛋白序列、16种Syt1蛋白序列、17种elav蛋白序列和2种Brp蛋白序列(序列信息详见表2)聚为4类,分别对应nSyb、Syt1、elav和Brp这4个基因家族(图1)。
单基因参考
Unigene reference基因名称
GeneBlastx最佳匹配(参考/名称/物种)
Blastx best hit (Reference/Name/Species)E值
E-valueInsectG05000 BdornSyb XM_011200958.4, vesicle-associated membrane protein 2 (Bactrocera dorsalis) 0 InsectG08551 BdorSyt1 XM_049446701.1, synaptotagmin 1 (B. dorsalis) 0 InsectG12478 Bdorelav XM_019990231.3, elav (B. dorsalis) 0 InsectG14028 BdorBrp XM_049452037.1, ELKS/Rab6-interacting/CAST family member 1 (B. dorsalis) 0 Table 1. nSyb, Syt1, elav, Brp genes identified in B. dorsalis
基因名称
Gene序列号
Sequence number缩写
AbbreviationDrosophila melanogaster bruchpilot isoform N UYI57892.1 DmBrp.N Bactrocera dorsalis bruchpilot BdorBrp Spodoptera littoralis Bruchpilot ASO76502.1 SlBrp Drosophila montana nSyb isoform X3 XP_064541710.1 DmonSyb.X3 Drosophila sulfurigaster albostrigata nSyb isoform X2 XP_062130853.1 DsulnSyb.X2 Drosophila nasuta nSyb isoform X2 XP_060658090.1 DnasnSyb.X2 Drosophila albomicans nSyb XP_034107380.1 DalbnSyb Drosophila melanogaster nSyb isoform G NP_ 728645.1 DmnSyb Drosophila suzukii nSyb XP_016933912.1 DsuznSyb Drosophila gunungcola nSyb isoform X1 XP_052847459.1 DgunnSyb.X1 Stomoxys calcitrans nSyb isoform X5 XP_013113466.1 ScalnSyb.X5 Musca domestica nSyb isoform X1 XP_058983425.1 MdomnSyb.X1 Musca vetustissima nSyb XP_061396193.1 MvetnSyb Bactrocera dorsalis nSyb BdornSyb Zeugodacus cucurbitae nSyb isoform X5 XP_011196106.1 ZcucnSyb.X5 Anastrepha obliqua nSyb isoform X3 XP_054734036.1 AoblnSyb.X3 Anastrepha ludens nSyb isoform X1 XP_053953598.1 AludnSyb.X1 Drosophila melanogaster synaptotagmin 1 isoform A NP_ 523460.2 DmSyt1.A Drosophila suzukii synaptotagmin 1 isoform X1 XP_016937535.1 DsuznSyt1.X1 Drosophila biarmipes synaptotagmin 1 isoform X1 XP_016954801.1 DbianSyt1.X1 Drosophila yakuba synaptotagmin 1 isoform X1 XP 002087728.1 DyaknSyt1.X1 Drosophila rhopaloa synaptotagmin 1 isoform X5 XP_016991250.1 DrhonSyt1.X5 Scaptodrosophila lebanonensis synaptotagmin 1 isoform X1 XP_030383437.1 SlebSyt1.X1 Drosophila busckii synaptotagmin 1 isoform X1 XP_017854866.1 DbusSyt1.X1 Calliphora vicina synaptotagmin 1isoform X4 XP_065357607.1 CvicSyt1.X4 Ceratitis capitata synaptotagmin1 isoform X1 XP_004537793.1 CcapSyt1.X1 Rhagoletis zephyria synaptotagmin 1 isoform X1 XP_017494880.1 RzepSyt1.X1 Anastrepha ludens synaptotagmin 1 isoform X4 XP_053961234.1 AluSyt1.X4 Anastrepha obliqua synaptotagmin 1 isoform X1 XP_054742113.1 AoblSyt1.X1 Zeugodacus cucurbitae synaptotagmin 1 isoform X1 XP_011192440.1 ZcucSyt1.X1 Bactrocera oleae synaptotagmin 1 isoform X4 XP_036230797.1 BoleSyt1.X4 Bactrocera oleae synaptotagmin 1 isoform X3 XP_014103290.1 BoleSyt1.X3 Bactrocera latifrons synaptotagmin 1 isoform X1 XP_018789563.1 BlatSyt1.X1 Bactrocera dorsalis synaptotagmin 1 BdorSyt1 Rhagoletis zephyria elav XP_017491017.1 Rzepelav Anastrepha ludens elav XP_053952087.1 Aludelav Anastrepha obliqua elav isoform X2 XP_054729194.1 Aoblelav.X2 Ceratitis capitata elav isoform X2 XP_004519191.1 Ccapelav.X2 Zeugodacus cucurbitae elav isoform X3 XP_028893842.1 Zcucelav.X3 Bactrocera oleae elav isoform X1 XP_014097934.1 Boleelav.X1 Bactrocera latifrons elav isoform X1 XP_018782793.1 Blatelav.X1 Bactrocera dorsalis elav Bdorelav Lucilia cuprina elav KAI8116666.1 Lcupelav Calliphora vicina elav XP_065364064.1 Cvicelav Drosophila melanogaster elav isoform B NP_001014713.1 Dmelav Drosophila obscura elav XP_041450494.1 Dobselav Drosophila virilis elav EDW63036.2 Dvirelav Drosophila innubila elav XP_034490791.1 Dinnelav Drosophila novamexicana elav XP_030568764.1 Dnovelav Drosophila montana elav isoform X2 XP_064547165.1 Dmonelav.X2 Drosophila mojavensis elav isoform X2 XP_032587857.1 Dmojelav.X2 Drosophila navojoa elav isoform X1 XP_017963840.1 Dnavelav.x1 Table 2. Sequence numbers used in conserved motif analysis
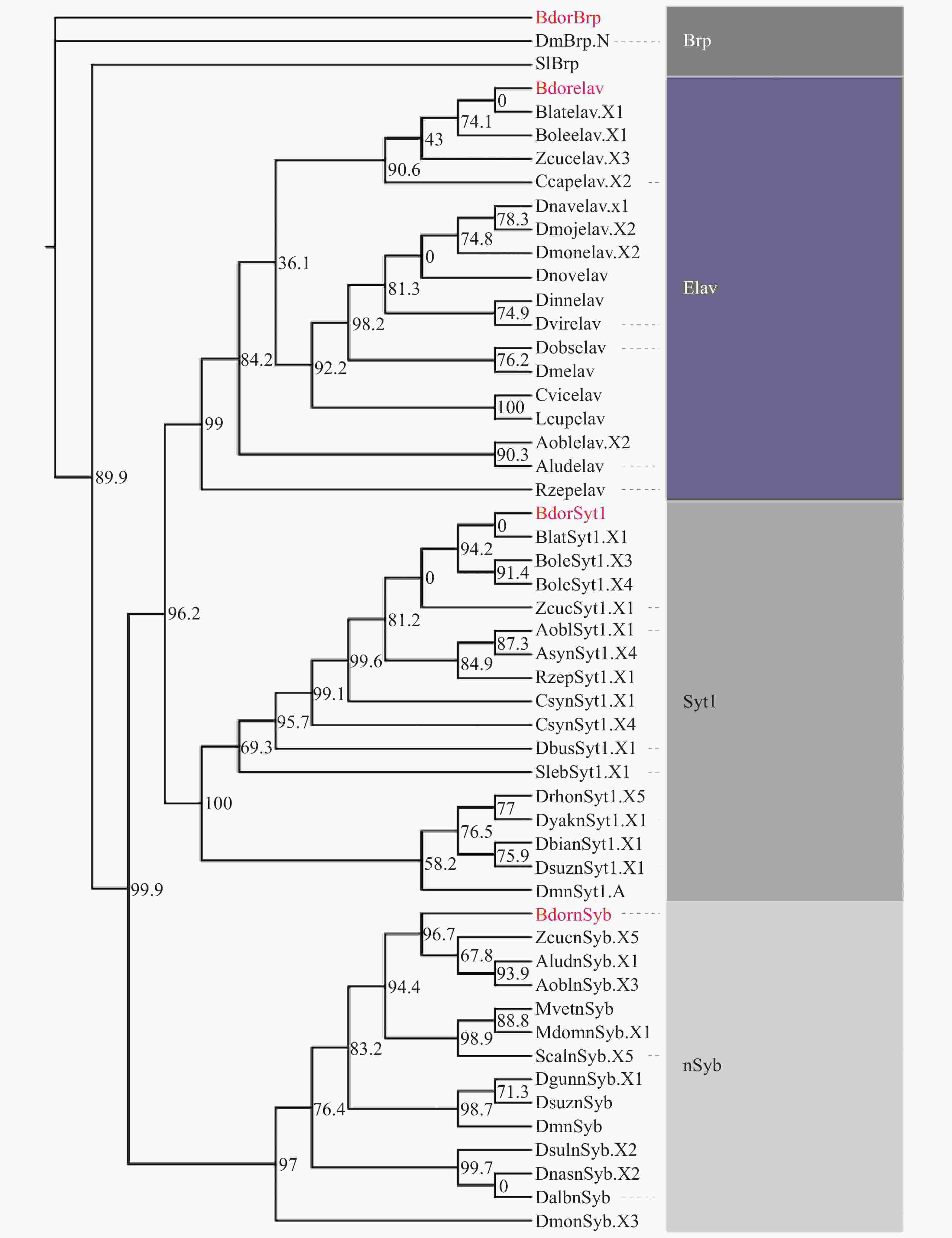
Figure 1. Phylogenetic analyses of BdornSyb, BdorSyt1, Bdorelav and BdorBrp
-
在上述橘小实蝇4种泛神经元表达基因鉴定的基础上,进一步结合系统发育树构建选取基因对应的蛋白序列进行保守基序分析(图2)。结果表明,nSyb蛋白共包含8个保守基序,不同物种的蛋白序列包括的保守基序数量为4~7个。除西印度按实蝇(Anastrepha obliqua)外,所有序列均含有5个相同的保守基序,顺序为“2-1-3-5-6”,基序8只存在于橘小实蝇和西印度按实蝇中,其中,橘小实蝇nSyb还缺乏保守基序4(图2-A);Syt1蛋白共包含10个保守基序,不同物种的蛋白序列包含的保守基序数量为9~10个。其中,黎巴嫩花果蝇(Scaptodrosophila lebanonensis)、醋果蝇(Drosophila busckii)和红头丽蝇(Calliphora vicina )中无保守基序10。橘小实蝇Syt1蛋白和大多数物种序列一致,包含10个保守基序,顺序为“10-8-5-4-7-2-9-6-3-1”(图2-B);Elav蛋白序列共包含10个保守基序,不同物种的保守基序数量均为10个,橘小实蝇与其他实蝇的序列结构高度相似,顺序均为“10-8-1-4-9-6-2-7-5-3”(图2-C);Brp蛋白共包含8个保守基序,不同物种保守基序数量为1~7个(SIBrp仅包含2个,推测是序列的不完整性导致的)。橘小实蝇的Brp蛋白保守基序中无保守基序2,其他基序类型与果蝇高度相似,顺序为“1-5-6-8-3-4-7”(图2-D)。
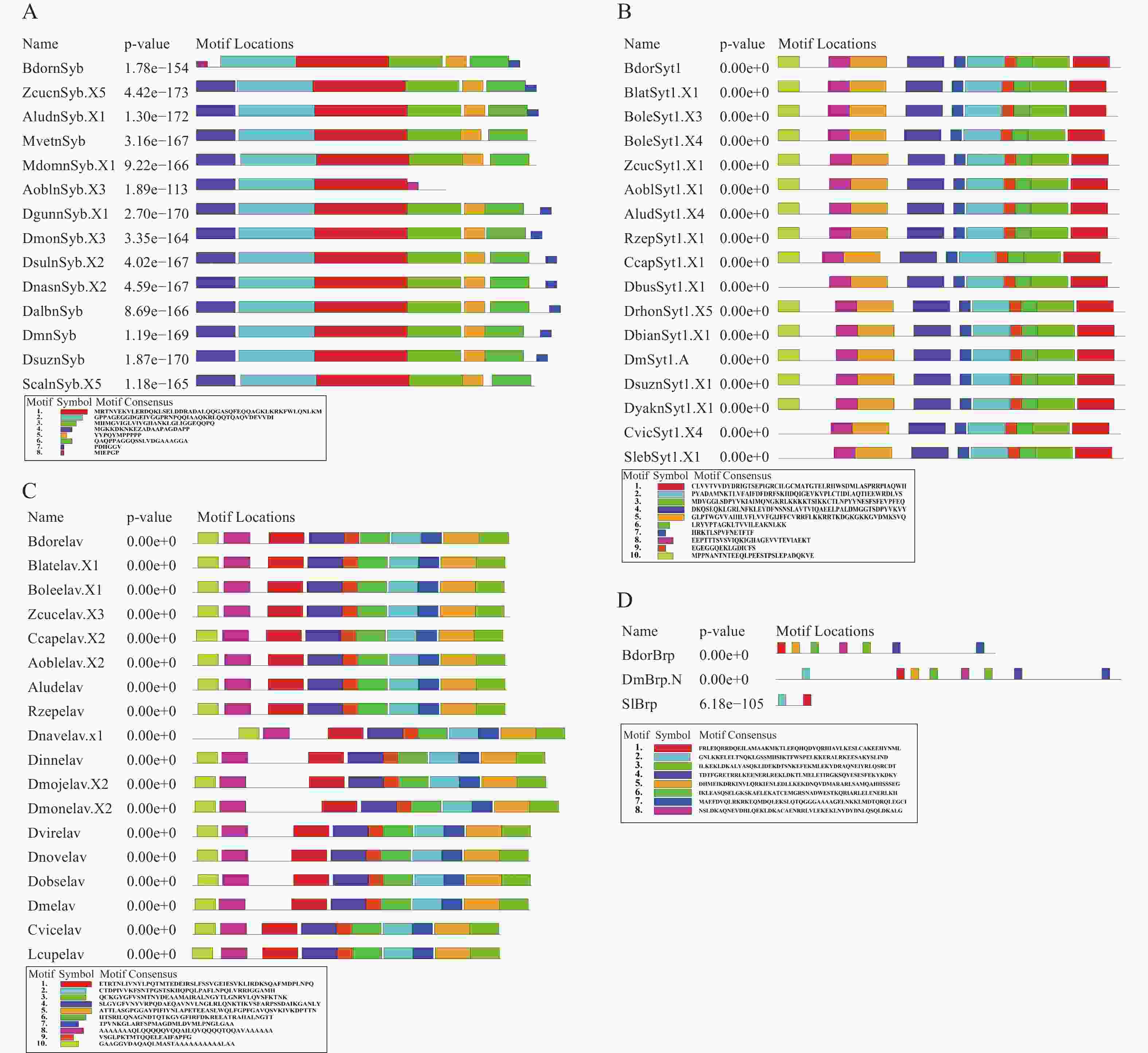
Figure 2. Conserved Motif analysis of BdornSyb, BdorSyt1, Bdorelav, and BdorBrp
-
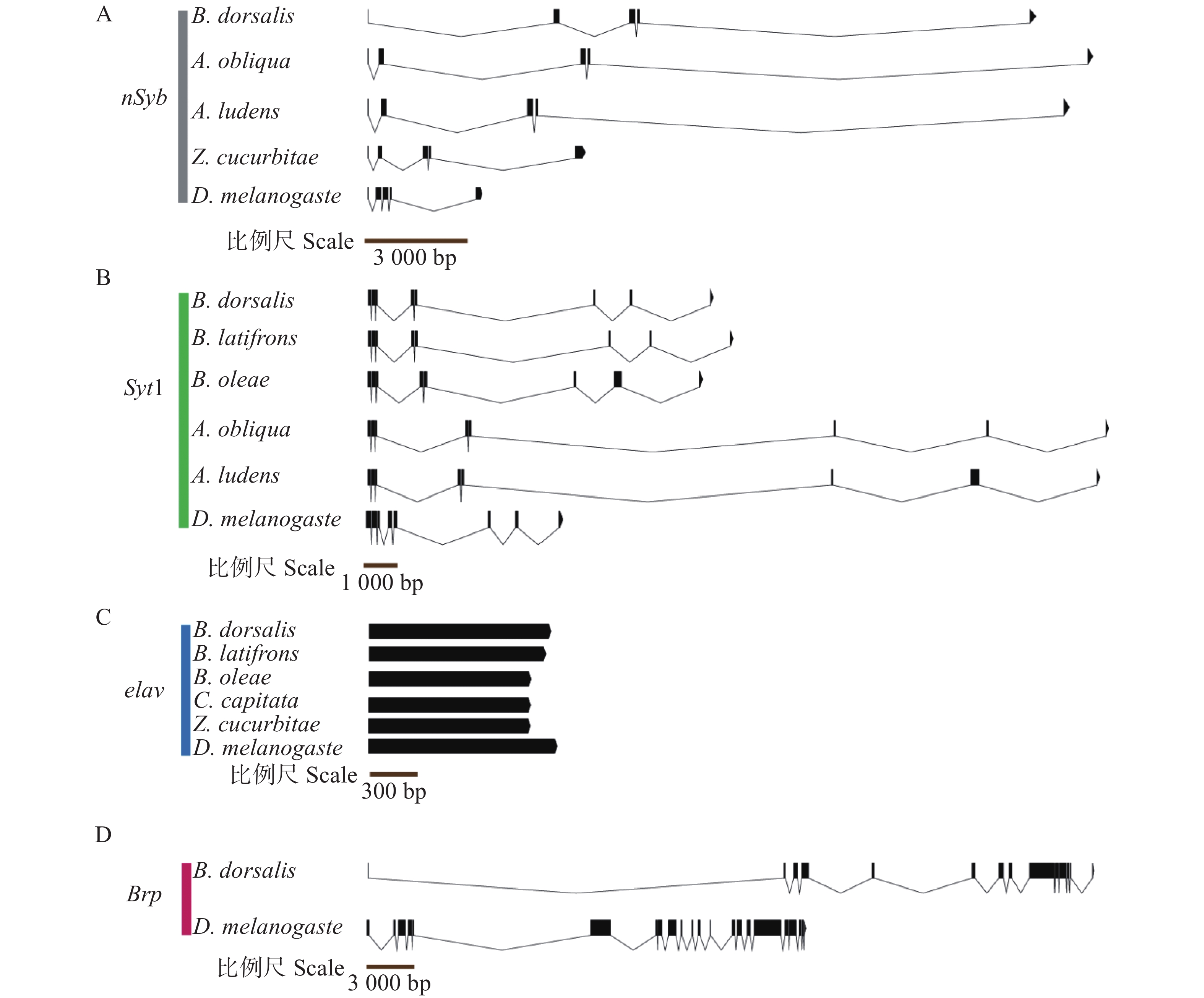
通过已有组装完整的转录组和基因组数据(国家生物信息中心 GSA 数据库,PRJCA020830),获得BdornSyb、BdorSyt1、Bdorelav、BdorBrp完整的外显子—内含子基因结构,并与黑腹果蝇及几种有报道结构的近缘实蝇科昆虫进行了比对(图3和表3)。结果表明,BdornSyb基因组长度为19 337 bp,包含5个外显子、4个内含子,与近缘种西印度按实蝇(Anastrepha obliqua)、墨西哥按实蝇(A. ludens)、瓜实蝇(Zeugodacus cucurbitae)以及黑腹果蝇(Drosophila melanogaster)具有相同数量的外显子数(图3-A)。BdorSyt1基因组长度为26 884 bp,包含8个外显子、7个内含子,与近缘种辣椒实蝇(Bactrocera latifrons)、橄榄实蝇(B. oleae)、西印度按实蝇(A. obliqua)、墨西哥按实蝇(A. ludens)及黑腹果蝇(D. melanogaster)具有相同数量的外显子数(图3-B)。Bdorelav基因结构特殊,只包含1个外显子,无内含子,整个基因组长度为1 341 bp;辣椒实蝇(B. latifrons)、橄榄实蝇(B. oleae)、地中海实蝇(Ceratitis capitata)、瓜实蝇(Z. cucurbitae)以及黑腹果蝇(D. melanogaster)中的Elav基因结构与Bdorelav相似,也只有1个外显子(平均长度1 278.00~1 440.00 bp)(图3-C)。BdorBrp基因组长度为49 692 bp,包含14个外显子和13个内含子,而黑腹果蝇的Brp包含22个外显子和21个内含子(图3-D)。总体而言,除Brp外,另外3种基因的结构域在近缘种中都高度保守。
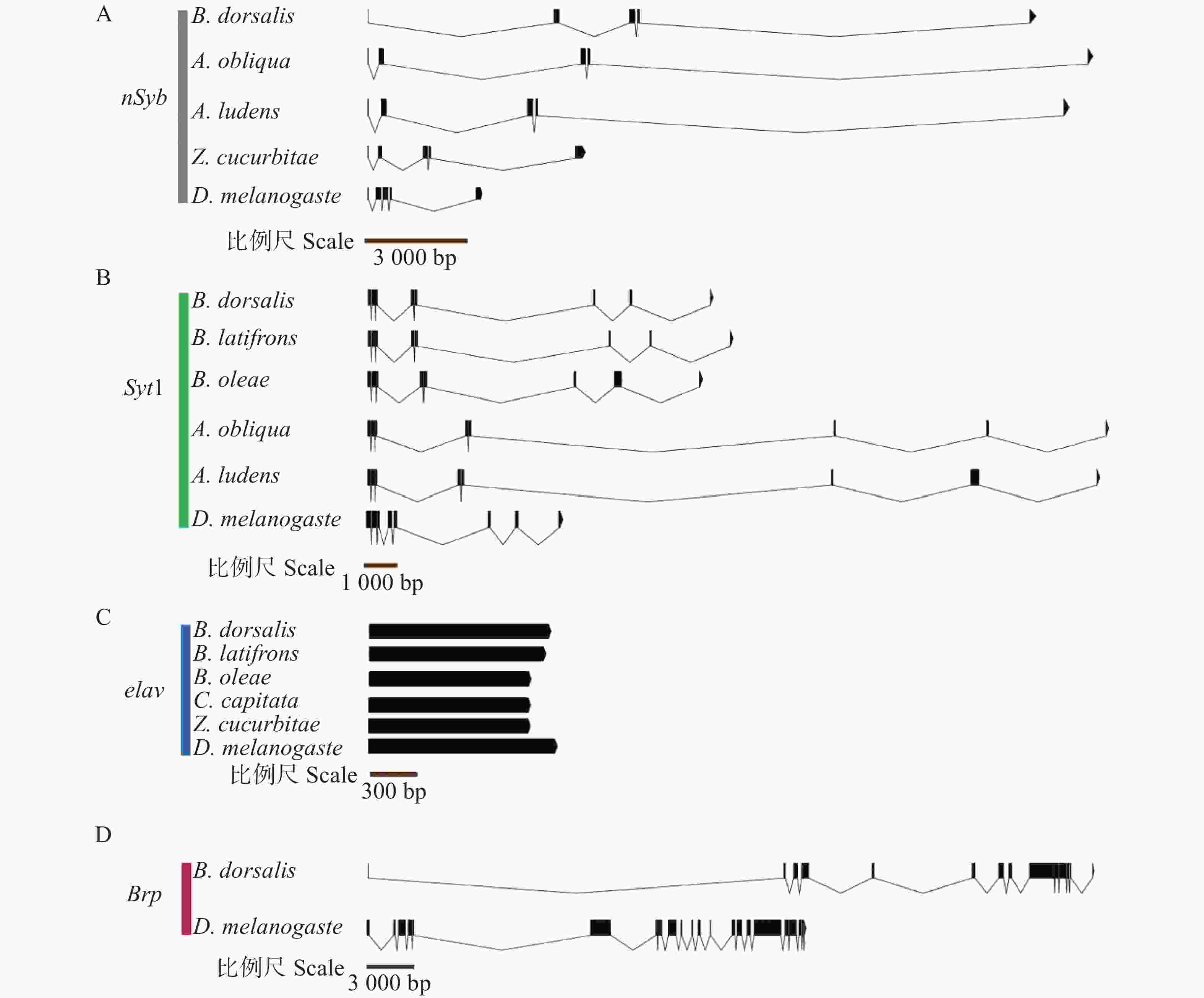
Figure 3. Exon-intron gene structures of BdornSyb, BdorSyt1, Bdorelav and BdorBrp
基因名称
Gene物种名称
Species外显子数量/个
Number of exons/ind外显子平均长度/bp
Average length
of exons/bp内含子数量/个
Number of
introns/ind内含子平均长度/bp
Average length
of introns/bpnSyb 橘小实蝇 Bactrocera dorsalis 5 105.60 4 4702.25 西印度按实蝇Anastrepha obliqua 5 111.60 4 6308.50 墨西哥按实蝇 A. ludens 5 111.60 4 5251.50 瓜实蝇 Zeugodacus cucurbitae 5 154.80 4 1887.25 黑腹果蝇 Drosophila melanogaster 5 115.80 4 780.00 Syt1 橘小实蝇 B. dorsalis 8 174.00 7 3641.71 辣椒实蝇 B. latifrons 8 174.00 7 3761.57 橄榄实蝇 B. oleae 8 216.00 7 2962.43 西印度按实蝇 A. obliqua 8 174.75 7 8321.00 墨西哥按实蝇 A. ludens 8 237.38 7 7890.71 黑腹果蝇 D. melanogaster 8 178.13 7 1342.29 elav 橘小实蝇 B. dorsalis 1 1341.00 0 0 辣椒实蝇 B. latifrons 1 1332.00 0 0 橄榄实蝇 B. oleae 1 1278.00 0 0 地中海实蝇 Ceratitis capitata 1 1278.00 0 0 瓜实蝇 Zeugodacus cucurbitae 1 1278.00 0 0 黑腹果蝇 D. melanogaster 1 1440.00 0 0 Brp 橘小实蝇 B. dorsalis 14 305.57 13 3493.38 黑腹果蝇 D. melanogaster 22 308.05 21 1060.95 Table 3. Number and average length of exons and introns of BdornSyb, BdorSyt1, Bdorelav, BdorBrp
-
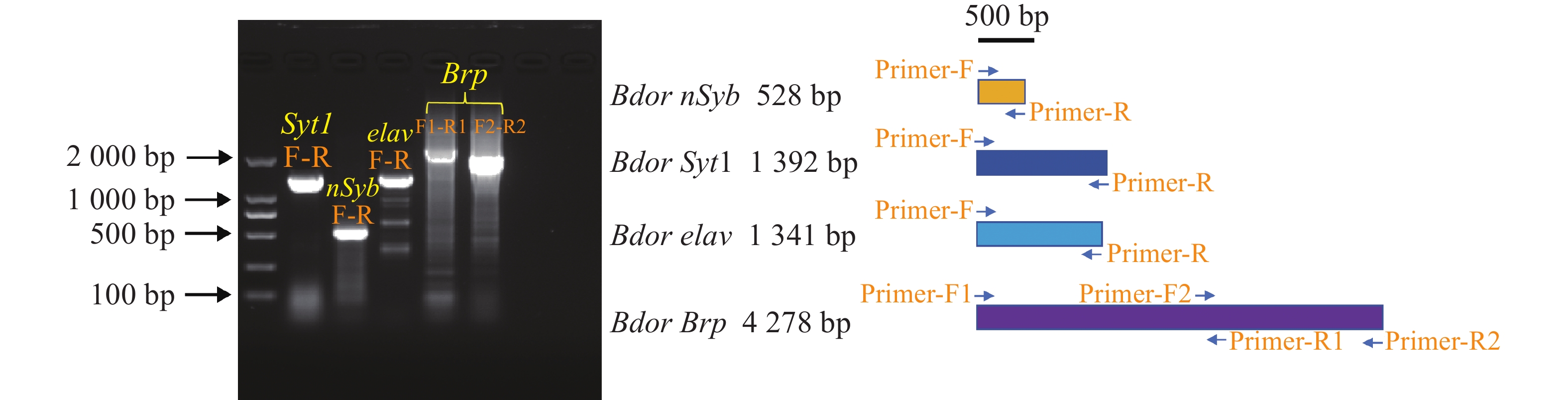
为确定序列的准确性,在基因结构的基础上,设计了特异性的扩增引物(引物和PCR条件详见表4)对CDS序列进行了验证,并成功克隆了四种基因的CDS序列,电泳胶图显示了与目标序列一致的长度(图4)。将条带回收后连接至blunt载体,进一步通过生工测序验证了目标条带的正确性。其中,BdornSyb CDS序列长528 bp,编码一个由175个氨基酸组成的蛋白,理论等电点为4.87,分子质量为18.57 kDa,不稳定性指数为51.33,脂肪族指数为78.69,平均亲水性指数为−0.509;BdornSyt1 CDS序列长1 392 bp,编码一个由463个氨基酸组成的蛋白,理论等电点为5.53,分子质量为52.39 kDa,不稳定性指数为43.26,脂肪族指数为81.02,平均亲水性指数为−0.462;Bdorelav CDS序列长1 341 bp,编码一个由446个氨基酸组成的蛋白,理论等电点为8.92,分子质量为47.18 kDa,不稳定性指数为38.77,脂肪族指数为79.15,平均亲水性指数为−0.208;BdorBrp CDS序列长4 278 bp,编码一个由1 425个氨基酸组成的蛋白,理论等电点为5.69,分子质量为16.65 kDa,不稳定性指数为56.39,脂肪族指数为73.03,平均亲水性指数为−2.201。

Figure 4. Gel images of PCR amplification and primer positions of CDS sequences of four genes
基因
GenePCR引物
PCR primersPCR条件
PCR conditionsBdornSyb F: ATGGCGGAACCAGCACC
R: TTATACTGCACCGTGCTGCTCT98 ℃ 3 min;98 ℃ 10 s,55 ℃ 15 s,72 ℃ 延伸 40 s,共40循环;72 ℃延伸10 min;10 ℃保存 BdorSyt1 F: ATGCCGCCAAACACGAATACG
R: TTACTTCATATTCTTCAGTATTTCATCGGT98 ℃ 3 min;98 ℃ 10 s,55 ℃ 15 s,72 ℃ 延伸 1 min 40 s,共40循环;72 ℃延伸10 min;10 ℃保存 Bdorelav F: ATGGACTTTATGATGGCGAATGC
R: TTACTTTGATTTGTTGGTCTTGAAGCT98 ℃ 3 min;98 ℃ 10 s,55 ℃ 15 s,72 ℃ 延伸 1 min 30 s,共40循环;72 ℃延伸10 min;10 ℃保存 BdorBrp F1: ATGGACAATGCCTACGTCTACTATAAGTT
R1: CGTTTCGAGTTCCATAAGGGTTTTGT
F2: GAGCCAAGTGGAGTTACGTAAACT
R2: TTAGAAAAAGCTCTTCAAGAATCCAGCTGGT98 ℃ 3 min;98 ℃ 10 s,55 ℃ 15 s,72 ℃ 延伸 2 min 30 s,共40循环;72 ℃延伸10 min;10 ℃保存 Table 4. Summary of primers and PCR conditions
-
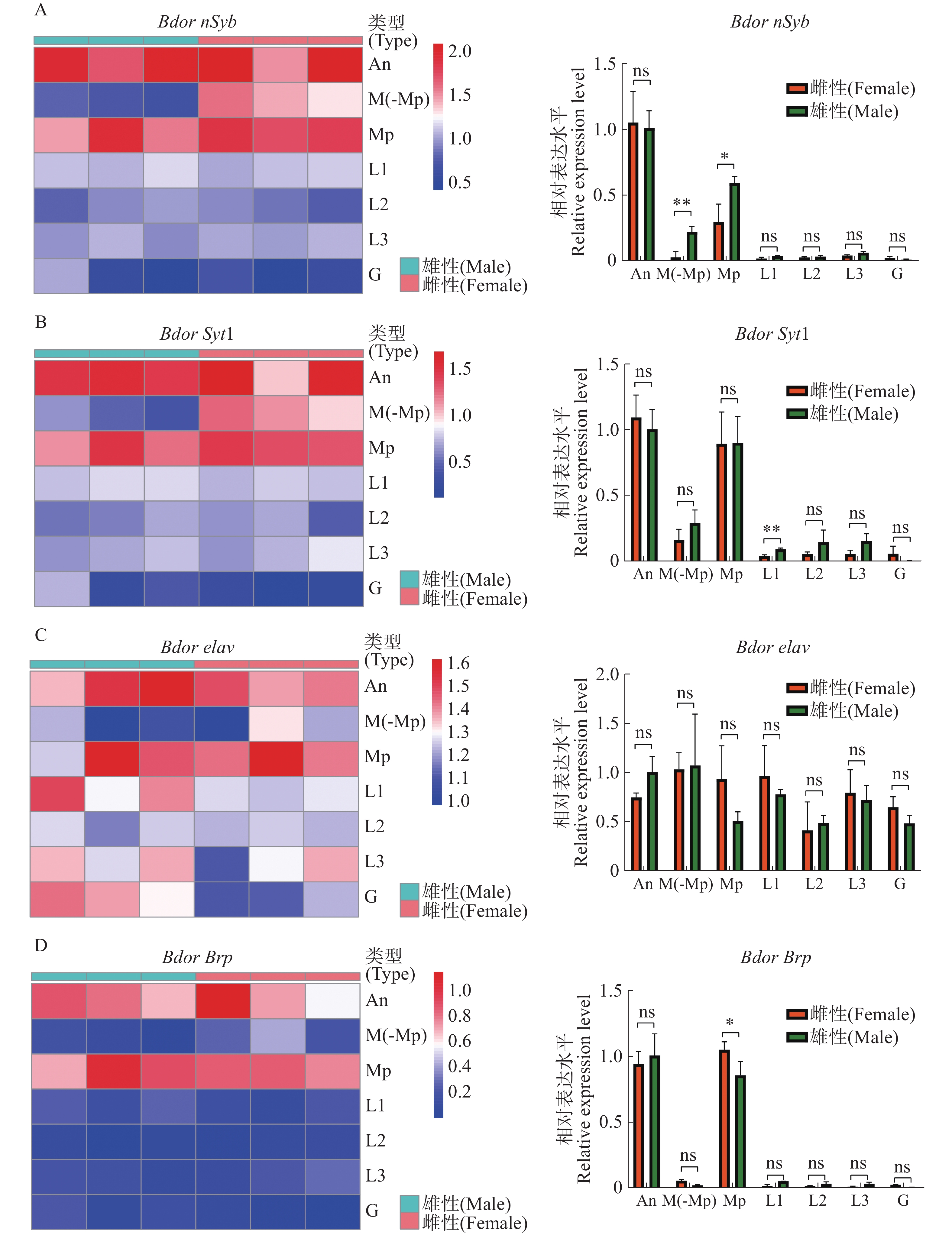
通过比较橘小实蝇雌雄外周感觉器官的转录组,获得了4种基因在外周的表达模式,并同时利用定量反转录聚合酶连锁反应(qRT-PCR)进行了验证。结果表明,BdornSyb和BdorSyt1在雌性触角和下颚须,雄性触角、口器(去下颚须)和下颚须中均显著表达,但在两性前、中、后足及外生殖器中的表达量较少(图5-A—B);雄性在口器(去下颚须)、下颚须上的BdornSyb表达量明显高于雌性,在前足上的BdorSyt1表达量显著高于雌性,其余组织雌雄表达量无显著差异。Bdorelav在雌雄中的表达模式没有明显差异,在各个组织均有一定的表达量,其中下颚须的表达量最高(图5-C);BdorBrp主要表达于触角和下颚须,其余组织几乎不表达,雌性下颚须上的表达量显著高于雄性(图5-D)。
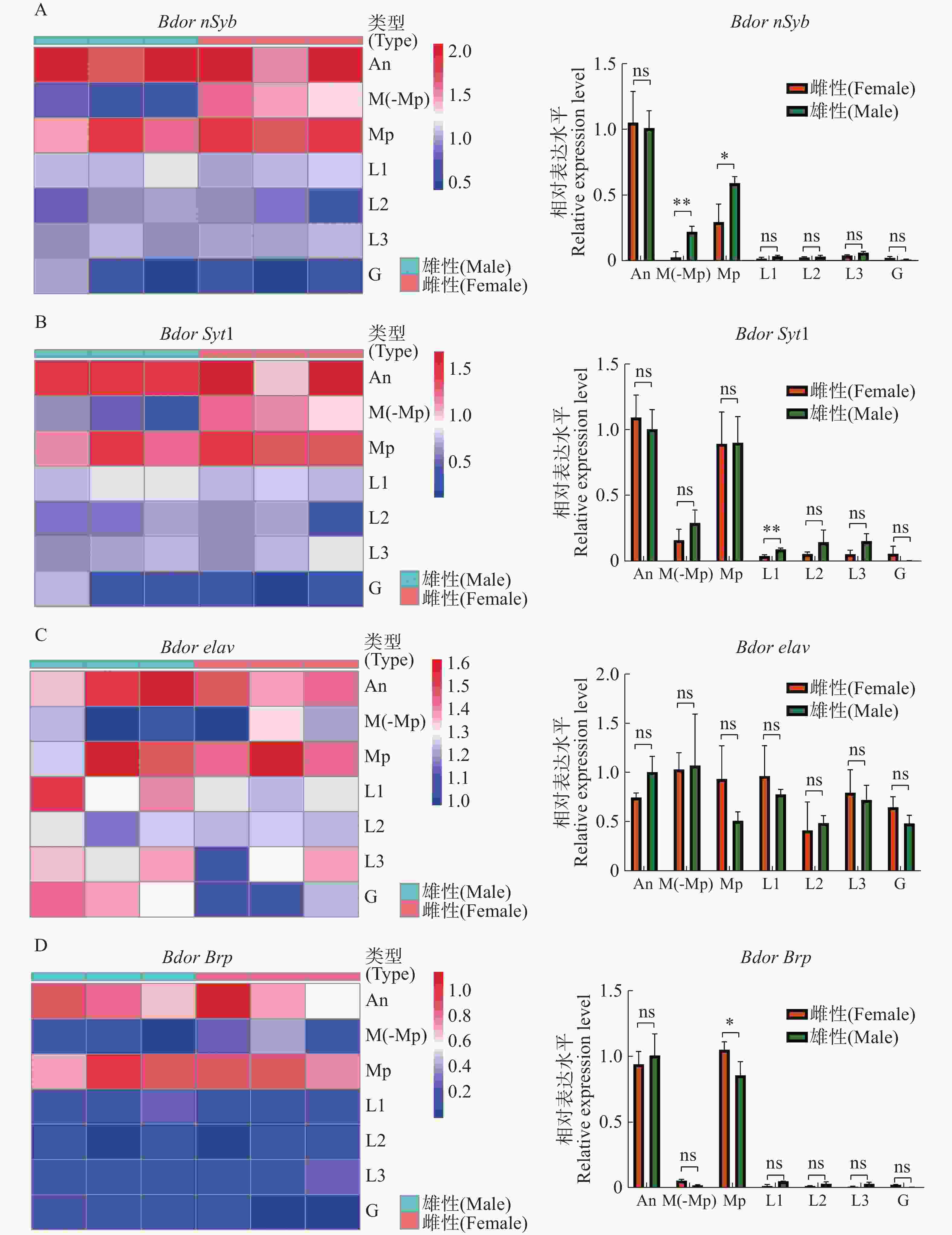
Figure 5. Peripheral nerve tissue expression profile of BdornSyb, BdorSyt1, Bdorelav and BdorBrp
-
中枢神经系统对害虫的行为,尤其是嗅觉行为,具有关键影响力。当外界化学信号被外周嗅觉神经转化为电信号后,这些信号通过神经传递至中枢神经系统进行整合和编码[21-22]。在这个过程中,突触在神经信号的传递、调节和整合中起到了至关重要的作用。当前昆虫中报道的泛神经元表达基因大多与突触功能相关,如Brp (Bruchpilot)、nSyb (神经突触素b)和Syt1 (突触融合蛋白1)。这些基因与突触功能的紧密关联,反映了突触在神经信号传递中的关键性。通过与果蝇进行同源比对,本研究鉴定出橘小实蝇的泛神经元表达候选基因。在近缘物种中,高度同源的基因通常表现出相似的基因结构特征[45-46]。本研究涉及的这些基因结构在不同的双翅目昆虫中相对保守,暗示了突触传递方式在这一类昆虫中的保守性。虽然这些基因在双翅目昆虫中共享保守基序,但也存在一些差异。例如,BdornSyb缺乏保守基序4但具有大多数物种没有的基序8。除了基序的变化,不同基因的内含子平均长度也显示出明显差异。例如,nSyb的内含子平均长度为780.00~6 308.50 bp,而Syt1的内含子平均长度为1 342.30~8 321.00 bp。这些内含子长度的变化提示,在基因表达调控上可能存在物种特异性机制。这种变异可能影响基因的转录和翻译效率,进而影响突触功能及行为适应性。这些基因结构和内含子长度的多样性强调了即使在保守的神经传递机制中,仍有显著的遗传多样性。这种多样性可能为不同物种提供了适应环境变化和行为多样化的潜力,并且为探索昆虫嗅觉系统的进化和功能提供了线索。
在果蝇中,nSyb、Syt1、Elav和Brp几乎在所有类型的神经元中表达[26,47]。这些基因在昆虫中的各种感受器类型中都有表达。例如,在蚊子中,Brp驱动的报告基因能够表达于诸如触角、口器、足和产卵器等部位,涵盖了嗅觉、味觉及机械感觉等感受器[30]。在本研究中,这些神经广泛表达基因的相对表达量存在差异,这与已有报道的物种类似。在蚊子中,nSyb和Syt1的转录水平是Brp的30倍~60倍[26]。对于相同的基因,其在外周神经组织中的表达量也存在差异。对于BdornSyb、BdorSyt1和Bdorbrp,触角及口器中的表达较高,这可能与这些外周神经器官上的神经细胞数量有关。例如,在蚊子中,利用Brp可以在触角和口器中标记超过1 000个神经细胞,而在足部平均只能标记大约40个神经细胞[30]。相比本研究团队鉴定的与突触相关的广泛神经元表达基因,Bdorelav的表达模式差异较大,在外周感觉器官中有明显表达,尤其是在可能神经细胞较少的生殖器部位,暗示该基因可能还在非神经元细胞中表达。在果蝇中,非神经组织同样转录Elav,但在转录后水平上被抑制[48]。这些基因表达特征的差异为本研究团队在构建转基因体系时提供了重要的选择依据,以优化不同神经标记的应用。
构建神经元遗传操作系统是泛神经表达基因应用的一个重要目标。目前,在果蝇中,利用nSyb和Elav驱动调控元件进行表达是最为广泛使用的方式,这些方法构建的果蝇品系已被众多研究者采用[29,49-50]。在非模式昆虫中,利用这些广泛神经元基因构建的工具主要包括两种:首先是神经细胞的标记,例如,在蚊子中,通过Brp驱动报告基因(如mCD8:GFP)来标记嗅觉器官中的神经元[30];其次是神经元活性的监测,例如,在蚊子和蜜蜂中,利用Brp或Synapsin驱动钙离子指示剂GCaMPs,以监测气味化合物在触角叶等神经区域的激活位置[26,51]。橘小实蝇是一种重要的农业害虫,目前的行为调控技术对防控橘小实蝇具有良好应用前景。尽管大量嗅觉分子靶标已被鉴定,但仍然缺乏“化学物质—嗅觉受体神经元—嗅小球—行为”之间的对应关系,这限制了嗅觉分子靶标在诱剂研发中的作用[12]。本研究鉴定的4种泛神经组织表达基因,是神经元发挥功能的基础,被认为是几乎在所有神经元中表达[26],故推测,这些基因可以广泛应用于外周感器神经元以及脑部神经元的标记中。为了进一步提升基因工具的特异性,未来研究应继续挖掘那些在特定类型神经元中表达的基因。例如,可以考虑利用BdorOrco专门标记所有气味受体神经元。这种特异性标记的策略,将为开发针对橘小实蝇的遗传工具提供新的可能,助力揭示橘小实蝇嗅觉神经通路中的复杂机制,并促进行为调控的深入研究和实际应用。
-
本研究通过进化和基因组结构分析,确认了BdornSyb、BdorSyt1、Bdorelav、BdorBrp在双翅目昆虫(尤其是黑腹果蝇)中的高度保守性。本研究团队发现这些基因在外周感觉器官中广泛表达,尤其在触角和下颚须中表现显著。基于这些基因此前在黑腹果蝇中的功能研究,本研究团队推测这些基因可以应用于构建外周泛神经元驱动体系,这将有助于开发更为精准和有效的防控措施,并拓展农业害虫管理的策略。
Identification and analysis of four pan-neuronal expression genes in Bactrocera dorsalis
DOI: 10.15886/j.cnki.rdswxb.20240158
- Received Date: 2024-10-21
- Rev Recd Date: 2024-11-07
- Available Online: 2025-03-10
- Publish Date: 2026-03-25
-
Key words:
- central nervous system function of insect pests /
- pan-neuronal expression genes /
- neuron labeling /
- Bactrocera dorsalis /
- olfactory
Abstract: Research on the central nervous system function of pests forms the foundation for developing precise behavior control technologies. Despite the significant role of neuron-labeling techniques based on genetic manipulation in this field, such techniques remain relatively scarce for non-model insects, such as the oriental fruit fly, Bactrocera dorsalis. In this context an attempt was made to identify four pan-neuronal expression genes in B. dorsalis with a view to laying the groundwork for constructing a neuron-labeling system for this species. The genomic structures of the pan-neuronal expression genes in B. dorsalis were identified and analyzed by employing bioinformatics and molecular biology to verify their full-length sequences and peripheral expression patterns. The results indicate that, by referring to four pan-neuronal expression genes from Drosophila, four homologous genes were identified in the B. dorsalis, namely BdornSyb, BdorSyt1, Bdorelav, and BdorBrp. The full genomic lengths of these four genes are 19,337 bp (5 exons, 4 introns), 26,884 bp (8 exons, 7 introns), 1,341 bp (1 exon), and 49,692 bp (14 exons, 13 introns), respectively. The domains of BdornSyb, BdorSyt1, and Bdorelav are highly conserved among closely related species. PCR cloning results indicated that the CDS sequence lengths of these four genes are all over 500 bp, consistent with the bioinformatics analysis results. Evolutionary and genomic structure analyses demonstrated that the four genes are highly conserved among Diptera insects. Expression pattern analysis revealed that all the four genes are expressed in the peripheral sensory organs of B. dorsalis, with three genes, BdornSyb, BdorSyt1 and BdorBrp, showing higher expression levels in the primary olfactory organs, the antennae, and the maxillary palp. The four genes identified are candidate pan-neuronal expression genes in B. dorsalis, providing a foundation for constructing a pan-neuronal labeling system for this species in the future.
| Citation: | Liu Jiaying, Zhang Jie, Wang Qi, Wu Shaoying, Wang Guirong, Liu Wei. Identification and analysis of four pan-neuronal expression genes in Bactrocera dorsalis[J]. Journal of Tropical Biology, 2026, 17(2): 233-246. doi: 10.15886/j.cnki.rdswxb.20240158

|
 DownLoad:
DownLoad: