
-
木薯(Manihot esculenta)是多年生灌木植物,广泛种植于热带和部分亚热带地区[1]。木薯具有耐旱、耐贫瘠、易栽培、块根富含淀粉等优良特性[2],一株木薯能够形成多达14个地下贮藏根,为约8亿人提供营养[3]。除了食用,木薯还用于制药、纺织、造纸和生物燃料等行业,具有巨大的经济价值[4]。但是木薯的病害会对其产量造成严重的损失,其中木薯细菌性枯萎病最为严重,它是由地毯草黄单孢杆菌木薯萎蔫致病变种Xanthomonas axonopodis pv. Manihotis(Xam)所引起的,会导致叶片枯萎和块根淀粉含量下降,严重时导致全株死亡[5]。因此寻求有效防控木薯细菌性枯萎病的方法是现在亟待解决的问题,分离鉴定木薯抗病基因是其中重要的一环[6]。
在病原菌长期侵害植物的过程中,植物形成了先天免疫系统[7],主要有病原相关分子模式激发的免疫反应(PAMP-triggered immunity,PTI)和效应蛋白激发的免疫反应(Effector-triggered immunity,ETI),且PTI和ETI二者协同互作[8],其中ETI是植物免疫系统中的一个重要部分,主要依赖于植物一类胞内免疫受体(Nucleotide-binding domain and leucine-rich repeat,NLR)识别病原微生物,调节植物免疫反应[9],该蛋白具有核苷酸结合域并富含亮氨酸重复序列(Nucleotide binding site-leucine-rich repeat,NBS-LRR)且高度保守[10],NLR蛋白主要包括3部分[11]:(1)可变的N端,能够激发下游传递免疫信号[12];(2)Nucleotide-binding adaptor shared by Apaf1,certain R genes and CED4(NB-ARC)结构域,可以与不同的核苷酸结合使NLR蛋白处于不同的状态[13];(3)C端的富亮氨酸重复类受体蛋白激酶(Leucine-rich-repeat,LRR)结构域,可以负调控NLR蛋白的活性[14]。
NLR是植物中非常重要的一类抗病基因家族(R基因),NLR广泛参与植物的生物胁迫[15]。有研究表明,在野生稻(Oryza officinalis)中含有NB-ARC结构域的基因是具有抗病功能的[16]。马铃薯(Solanum tuberosum)中的RySTO可编码NLR蛋白,转基因RySTO马铃薯植株可增强马铃薯对马铃薯疫病的抗性[17]。过表达TaRGA5基因,可增强小麦(Triticum aestivum)对白粉病的抗性[18],并且NLR正调控番茄对晚疫病(Phytophthora infestans)的抗性[19]。而在玉米(Zea mays)中突变含NLR结构的基因,导致水杨酸(Salicylic acid,SA)合成信号受到阻碍,进而负调控玉米抗病性[20]。在不同的物种中NLR结构都与调控病害息息相关[21],在木薯中过表达具有NLR结构的MeRPPL1(单个等位基因)基因,对南非木薯花叶病毒具有直接抗性[22]。MeLRRs是通过调控水杨酸含量和活性氧的积累以及致病相关基因1(Pathogenesis-related protein 1,PR1)的表达量,从而正调控木薯细菌性枯萎病的抗病性[23]。抗病小体(Zygote arrest-1,ZAR1)是通过识别病原微生物分泌效应因子,触发植物细胞死亡,从而阻止病原菌的扩散以达到抗病功能[24]。木薯中具NBS-LRR结构的基因众多,现已鉴定出228个NBS-LRR型基因和99个部分NBS基因[25],而大部分具NBS-LRR结构的基因的功能还有待进一步研究。
基于前人研究[26]和木薯NLR基因组序列,本研究鉴定了1个木薯体内编码NLR蛋白的基因MeRXam2,克隆了全长序列。对该基因进行了生物信息学分析以及探讨了其在木薯被病原菌侵染后的表达水平,并通过外源喷施蛋白探索MeRXam2蛋白对病原菌的影响,旨在为木薯分子育种提供候选蛋白。
-
供试材料为‘华南124’木薯品种,将木薯的茎秆截成10~15 cm长短(保留2~3芽点),种植在装有混有营养土和蛭石的花盆中,茎秆于28~30 ℃、12 h黑暗和12 h光照交替的条件下生长。
-
通过数据库phytozome(
https://phytozome-next.jgi.doe.gov/blast-search )查找MeRXam2的CDS序列,用Primer 5.0软件设计引物,引物合成和测序委托华大基因公司完成,具体引物信息如表1所示。引物名称
Primer碱基序列(5′−3′)
Primer sequence(5′−3′)用途
UseQMeRXam2-F TATAATTCAAGCCATTC qRT-PCR QMeRXam2-R ATCCAACAAGTCATCTG qRT-PCR Elongation factor 1-alpha-F TAACCACCCTGGTCAGATTGGAA 内参基因 Elongation factor 1-alpha-R AACTTGGGCTCCTTCTCAAGCTCT 内参基因 MeRXam2-F ATGGCAGAGATAGCCATATCC MeRXam2扩增 MeRXam2-R CTATTTCTGGAACTCAGGTGA MeRXam2扩增 MeRXam2-pET32a-F GCCATGGCTGATATCGGATCCATGGCAGAGATAGCCATATCCC 载体构建 MeRXam2-pET32a-flag-R TTGTCGACGGAGCTCGAATTCCTTGTCGTCATCGTCTTTGTAG 载体构建 TCTTTCTGGAACTCAGGTGACCCA Table 1. Primer sequences
-
取木薯叶片0.2~0.5 g,用液氮研磨充分,采用CTAB提取法提取木薯RNA。用反转录试剂盒(11151ES0,YEASEN)将RNA反转录为cDNA,每个RNA样品取4 μL,依次加入反转录酶混合体系5 μL、水11 μL,混合均匀后,37 ℃反应5 min,85 ℃终止反应,RNA与cDNA存放在−80 ℃冰箱保存备用。
-
以cDNA作为模板,用FastPfu DNA Polymeraset扩增MeRxam2片段,琼脂糖凝胶电泳检测基因片段大小,将目的片段用胶回收试剂盒回收,将回收的片段与 pEASY-Blunt3 连接,并转化到DH5α感受态细胞中(ZC101-2,ZOMANBIO),然后挑选单菌落进行菌落PCR验证,PCR程序设置程序如下:预变性95 ℃,3 min、变性95 ℃,30″、退火55 ℃,3 min、延伸72 ℃,根据片段长度设置
1 000 bp·min−1、变性至延伸设置循环n≥32次,最终延伸72 ℃,5 min。若为阳性克隆,挑选单一明亮的单克隆菌株送华大公司测序,测序返回结果与网站上的序列进行比对,将MeRXam2序列一致的菌液保存。 -
蛋白质的理化性质通过在线软件(
https://web.expasy.org/protparam/ )进行分析;通过在线软件(https://www.uniprot.org/ )绘制蛋白质的二级结构;蛋白质的三级结构通过在线工具SWISS-MODEL(https://swissmodel.expasy.org/ )进行预测;磷酸化位点通过(https://services.healthtech.dtu.dk/services/NetPhos-3.1/ )进行预测,蛋白的跨膜结构相关信息通过(https://services.healthtech.dtu.dk/services/TMHMM-2.0/ )预测,构建进化树所需序列在NCBI网址(https://blast.ncbi.nlm.nih.gov/Blast.cgi )上下载,利用MEGA.11软件进行进化树的构建。 -
用Xam侵染木薯叶片组织,然后分别取0、1、3、6、12和24 h的样品提取RNA,反转录为cDNA,用实时荧光定量PCR检测MeRXam2的表达量,使用0.1 mL 96孔qPCR板白色(KG2554,KIRGEN),在实时荧光定量系统(CFX Connect™ Real-Time,Bio-RadLaboratories)中进行。
-
提取构建成功的MeRXam2-Blunt3载体质粒,以质粒为模板,扩增MeRXam2的片段,用限制性内切酶BamH Ⅰ和EcoR Ⅰ双酶切载体pET32a,用同源重组连接的方法将MeRXam2片段连接到已经线性化的pET32a 载体上,转化后,经过菌落PCR验证,挑选具有单一明亮条带的菌落,并用LB液体培养基培养,提取质粒,双酶切验证,符合预期大小,并送华大公司测序,若序列一致,即视为构建载体成功。
-
将构建成功的质粒MeRXam2-pET32a转入到BL21(DE3)感受态细胞(ZC121-2,ZOMANBIO),经PCR验证,挑选具有单一明亮的菌落进行后续的实验。将过夜的菌液加入新鲜含氨苄青霉素钠抗性的LB液体培养基中,使菌液的OD600值为0.1~0.2,放入摇床中培养(37 ℃,150 r·min−1),当OD600值达到0.6左右,加入IPTG至终浓度为1 mmol·L−1,继续放入摇床中(37 ℃,150 r·min−1)进行培养并诱导蛋白。与此同时,取0、1、3、6 h的蛋白样品,离心(12 000 r·min−1,10 min),弃上清,菌体用100 μL PBS(pH7.4)重悬,超声波破碎,离心(4 ℃,12 000 r·min−1,5 min),弃上清液,沉淀用PBS悬起,然后进行十二烷基硫酸钠聚丙烯酰胺凝胶电泳(Sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDS-PAGE)和蛋白质免疫印迹(Western blot)检测,并测定蛋白浓度。
-
细菌数的测定:木薯在接种Xam后喷施蛋白,于0、3、6 d后用打孔器取相同大小的叶片,用75%(v/v)的乙醇浸泡5 min,灭菌水清洗3次,吸水纸吸干叶片表面水分,用研磨器将叶片组织磨碎,加入灭菌水混匀,在超净工作台里进行梯度稀释,并点在不含抗性的固体的培养基中,倒置放入28 ℃恒温培养箱进行培养,第2天进行细菌数的统计。
-
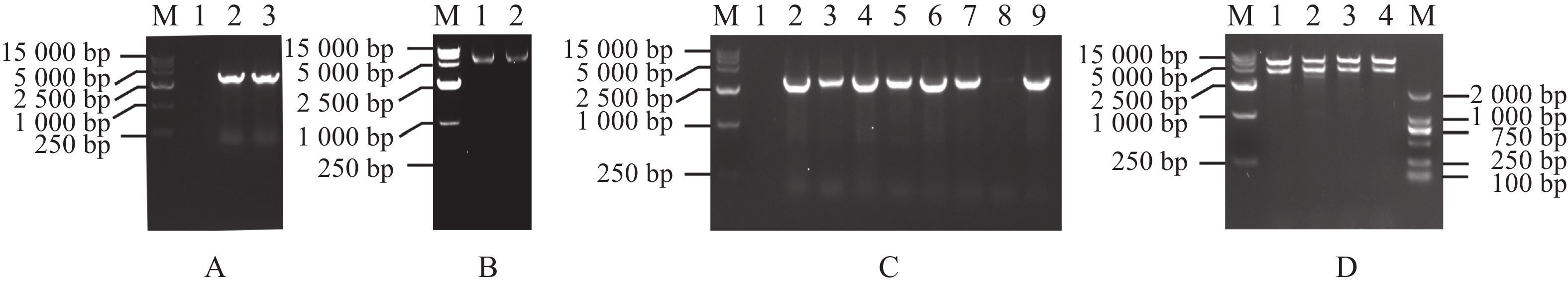
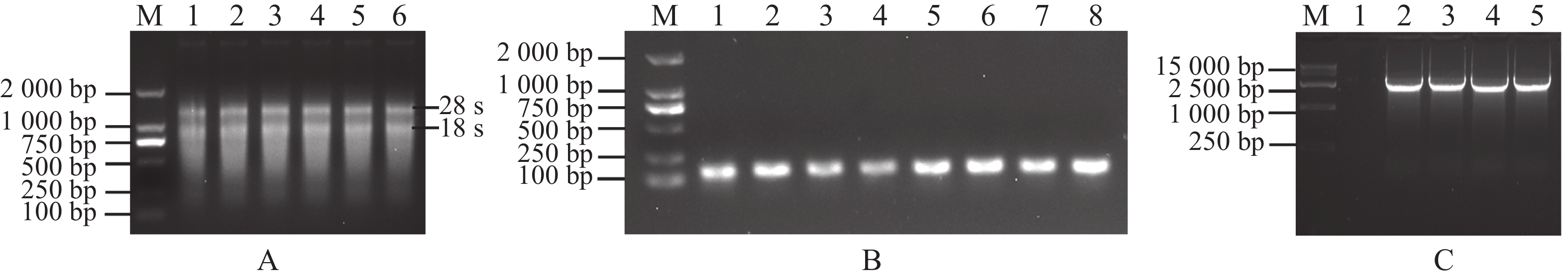
为克隆MeRXam2基因,首先提取木薯叶片总RNA,琼脂糖凝胶电泳检测RNA质量,发现有28、18 s条带(图1−A),即为RNA提取成功。然后将RNA反转录成cDNA,用木薯内参基因进行PCR检测cDNA的质量,发现可扩增单一条带,说明cDNA反转录成功(图1−B)。最后以cDNA为模板,扩增MeRXam2目的片段,得到与预测大小一样的片段(图1−C)。
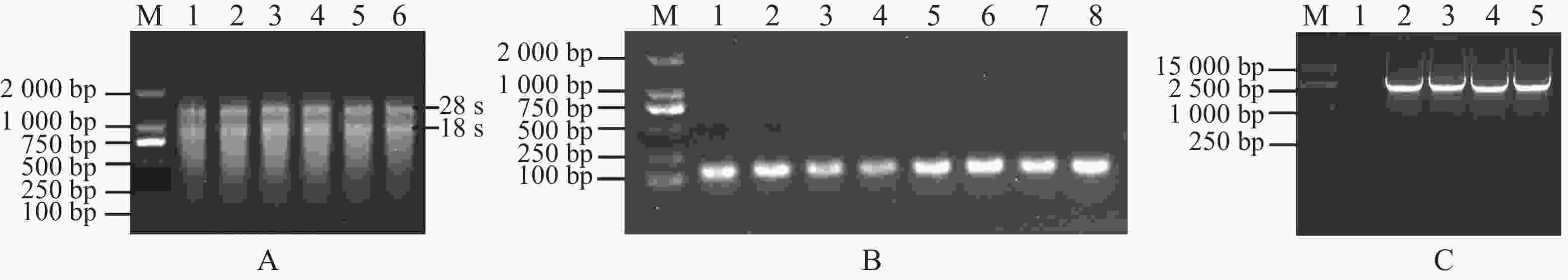
Figure 1. Acquisition of the MeRXam2 gene
-
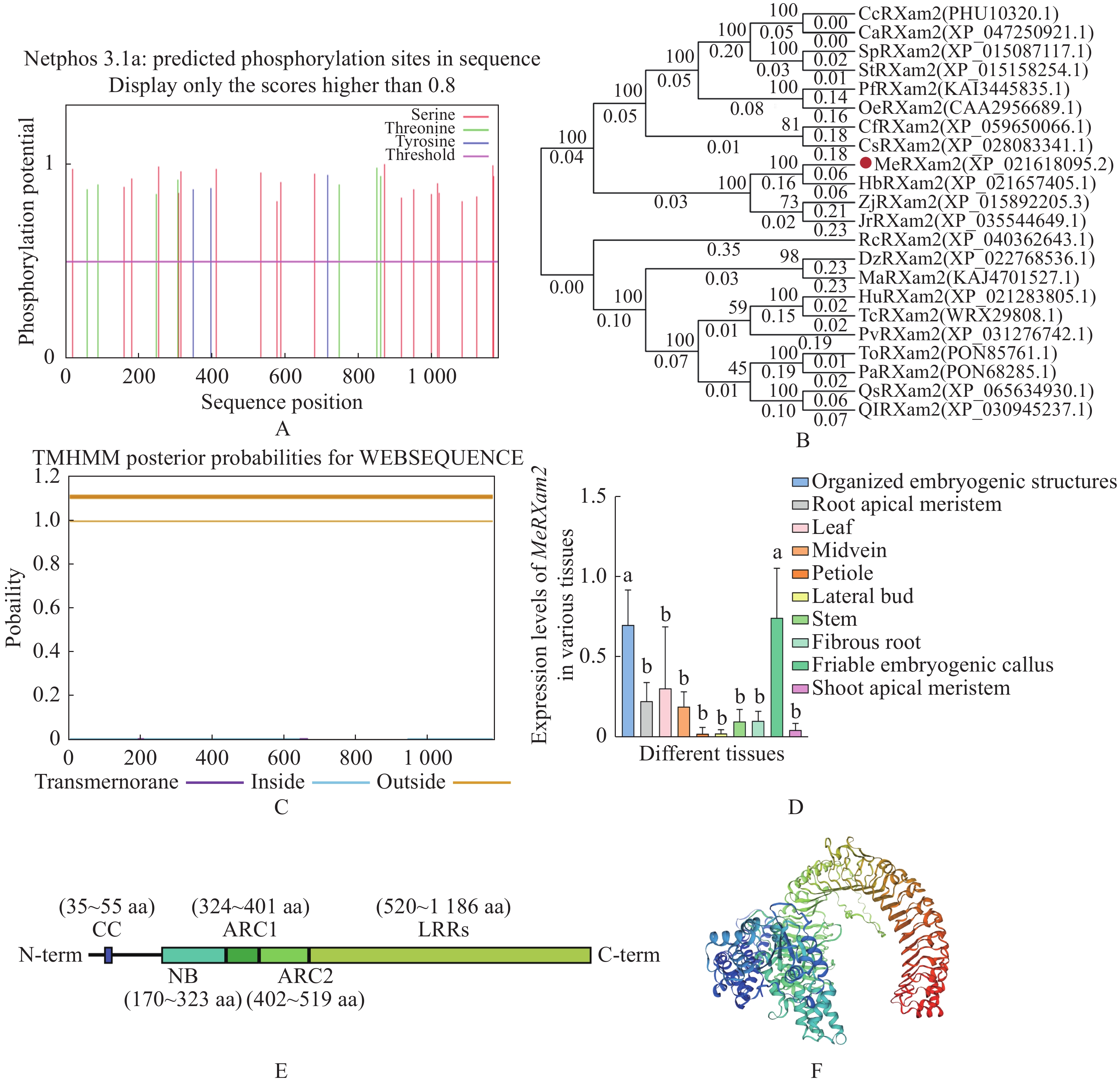
通过对MeRXam2蛋白磷酸化位点进行预测,发现有多个磷酸化位点(图2−A),通过构建进化树发现与MeRXam2同源性最高的是橡胶树(Hevea brasiliensis)HbRXam2 (图2−B),橡胶树和木薯同属于大戟科植物,两条蛋白序列同源性达到84.26%。此外,对蛋白进行了跨膜结构域预测,该蛋白不存在跨膜结构域(图2−C)。
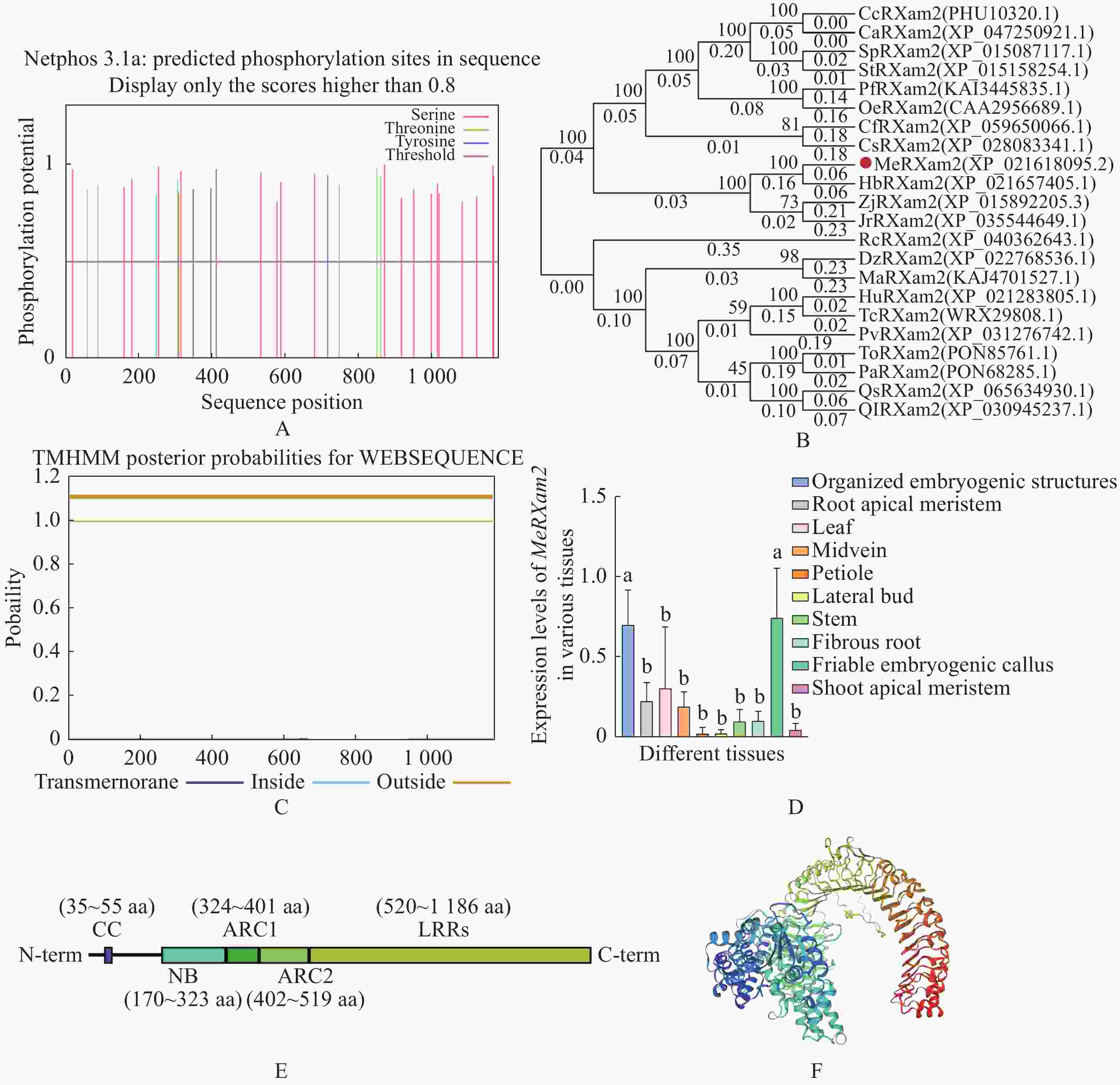
Figure 2. Physio-chemical analysis and evolutionary tree construction of MeRXam2 protein
基于TCOD expression在线数据库(
https://ngdc.cncb.ac.cn/tcod/expressions )中MeRXam2在不同组织的表达情况,并绘制成柱状图,发现MeRXam2在胚胎发生结构和易碎胚性愈伤组织的表达量相对较高(图2−D)。此外,本研究通过对蛋白结构的预测,发现MeRXam2结构主要包括3个区域:靠近N端的CC结构域(35~55 aa)、中间区域的NB-ARC(170~519 aa),NB-ARC其中又包含3个小区域,分别是NB结构域(170~323 aa),ARC1结构域(324~401 aa),ARC2结构域(402~519 aa)、还有靠近C端的LRRs结构域(520~1186 aa),LRRs是典型的NLR结构,根据在线软件(https://www.uniprot.org/ )提供的信息绘制了该蛋白的结构(图2−E)。另外,还通过网站绘制了MeRXam2的三级结构(图2−F)。蛋白质的理化性质通过在线软件(https://web.expasy.org/protparam/ )进行分析,结果表明,MeRXam2为木薯特异性免疫系统的受体NLR蛋白,CDS序列有3516 bp,氨基酸序列有1 186 aa,该蛋白分子式为C5947 H9596 N1598O1713 S61,预期蛋白大小约为132.847 kDa,理论等电点8.12。 -
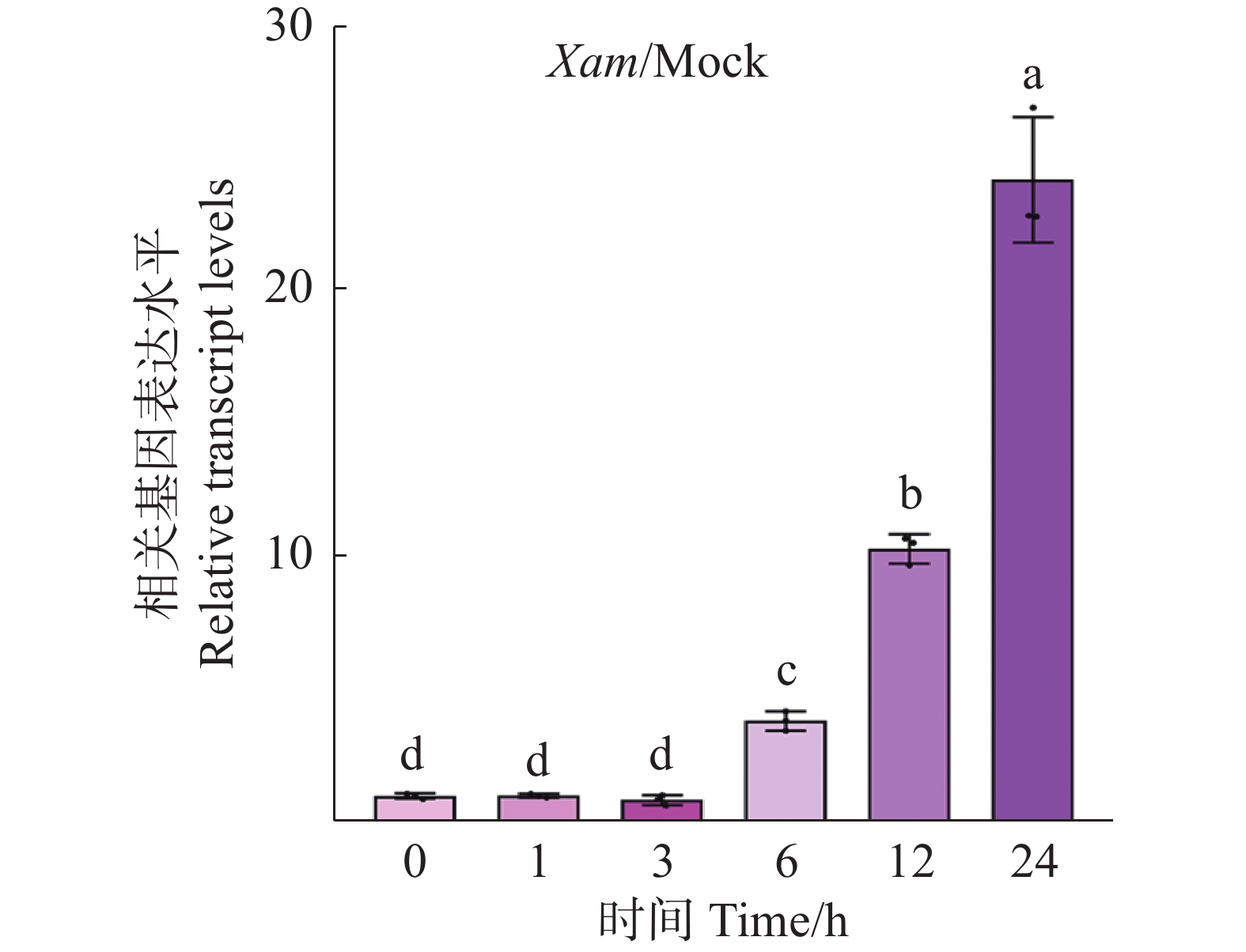
为了解MeRXam2是否参与到木薯的抗病过程中,采用实时荧光定量PCR技术检测MeRXam2基因在木薯被Xam处理0、1、3、 6、12和24 h后的表达水平。研究结果发现,随着时间的变化,MeRXam2表达量逐渐增加,且在24 h时的表达量达到最高(图3),这表明MeRXam2可能参与到木薯抗病反应过程中。
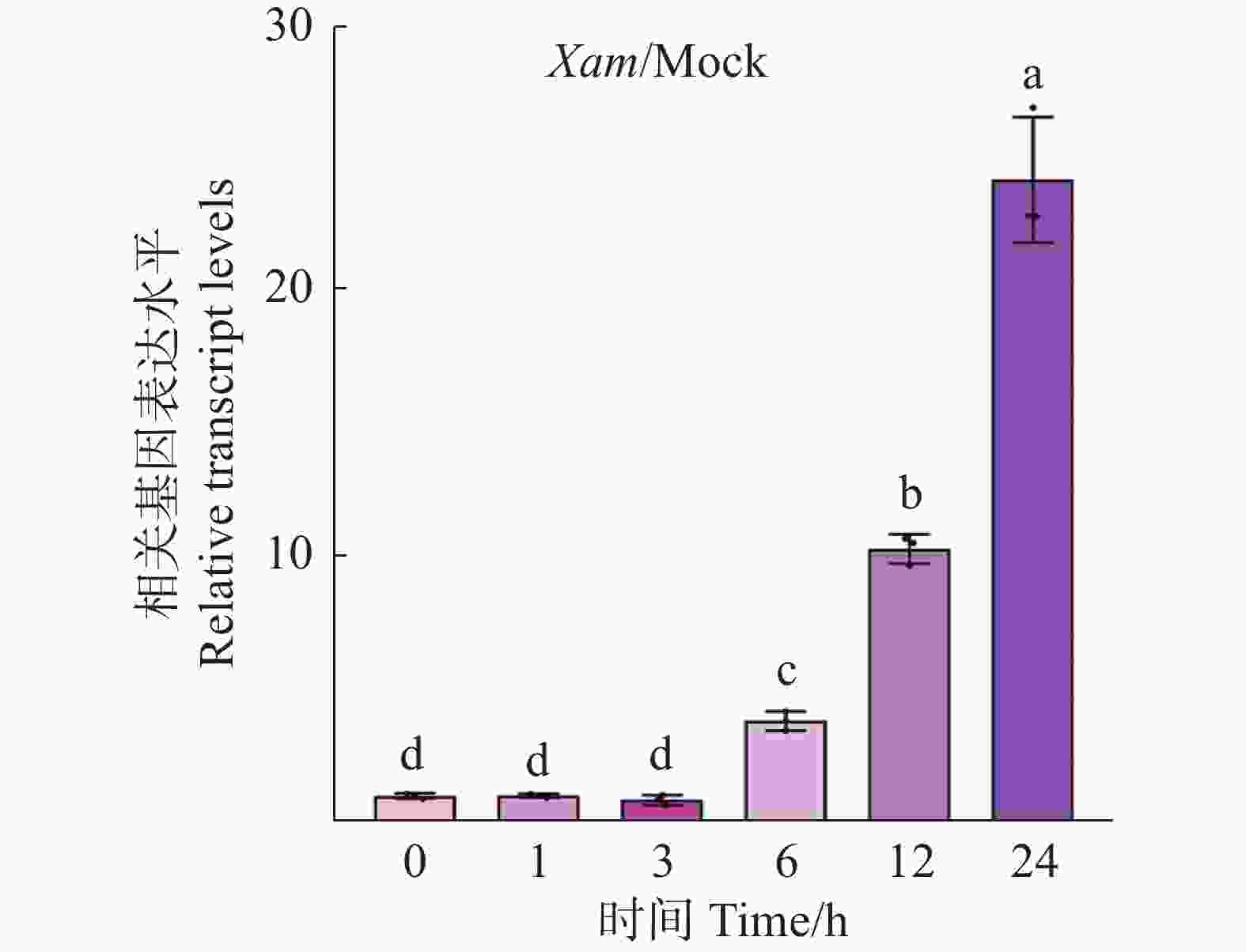
Figure 3. MeRXam2 gene expression profile level
-
为构建MeRXam2-pET32a原核蛋白表达载体,首先用PCR体系扩增了MeRXam2片段(图4−A),通过同源重组连接的方法,将目的基因与已用限制性内切酶BamH Ⅰ和EcoR Ⅰ双酶切好的载体pET32a连接(图4−B),转化到DH5α感受态细胞中,通过菌落PCR验证,挑选阳性单克隆(图4−C),用含氨苄青霉素钠抗性的液体LB培养,提取质粒,双酶切验证,切下来的目的片段大小符合(图4−D),并送测序验证,测序结果返回与序列一致,即为载体构建完成。
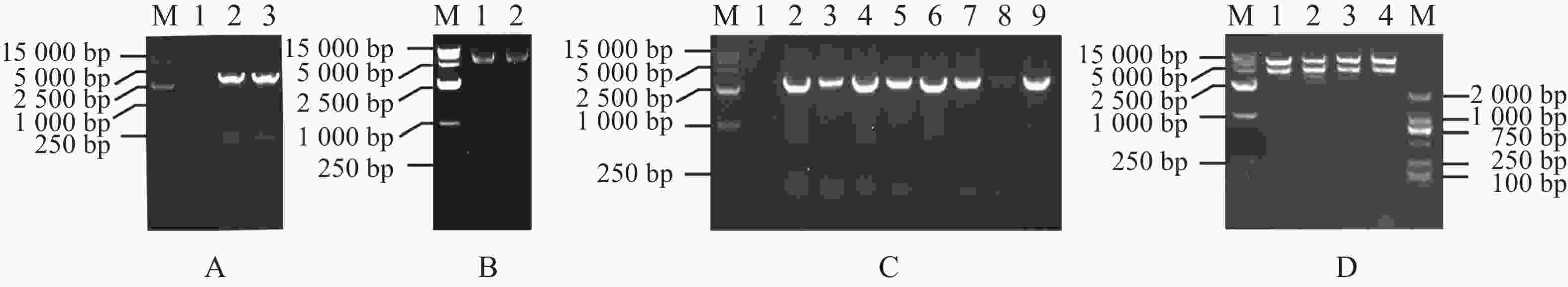
Figure 4. Construction of MeRXam2 gene vector
-
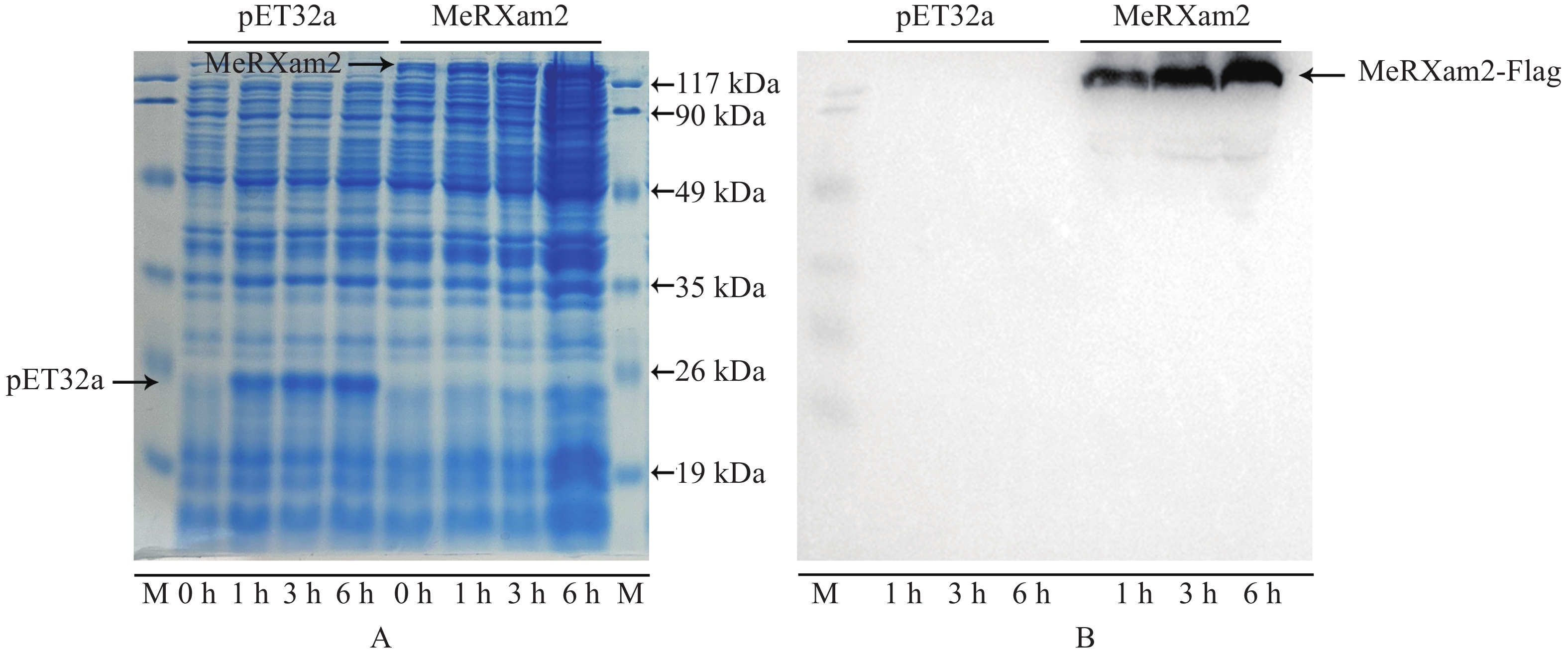
为探究蛋白的功能,本研究进行了MeRXam2-pET32a的蛋白诱导,结果发现能在37 ℃下诱导出蛋白,并且随着时间的增加,蛋白的浓度也增加(图5−A—图5−B)。为进行后续的实验,对6 h的蛋白用酶标仪测定蛋白浓度,MeRXam2蛋白在595 nm吸光度平均值为0.735,标准曲线Y=
0.5493 X+0.4016 (R2=0.993 8),最终得出蛋白质量浓度为0.608 g·L−1。
Figure 5. MeRXam2 protein
-
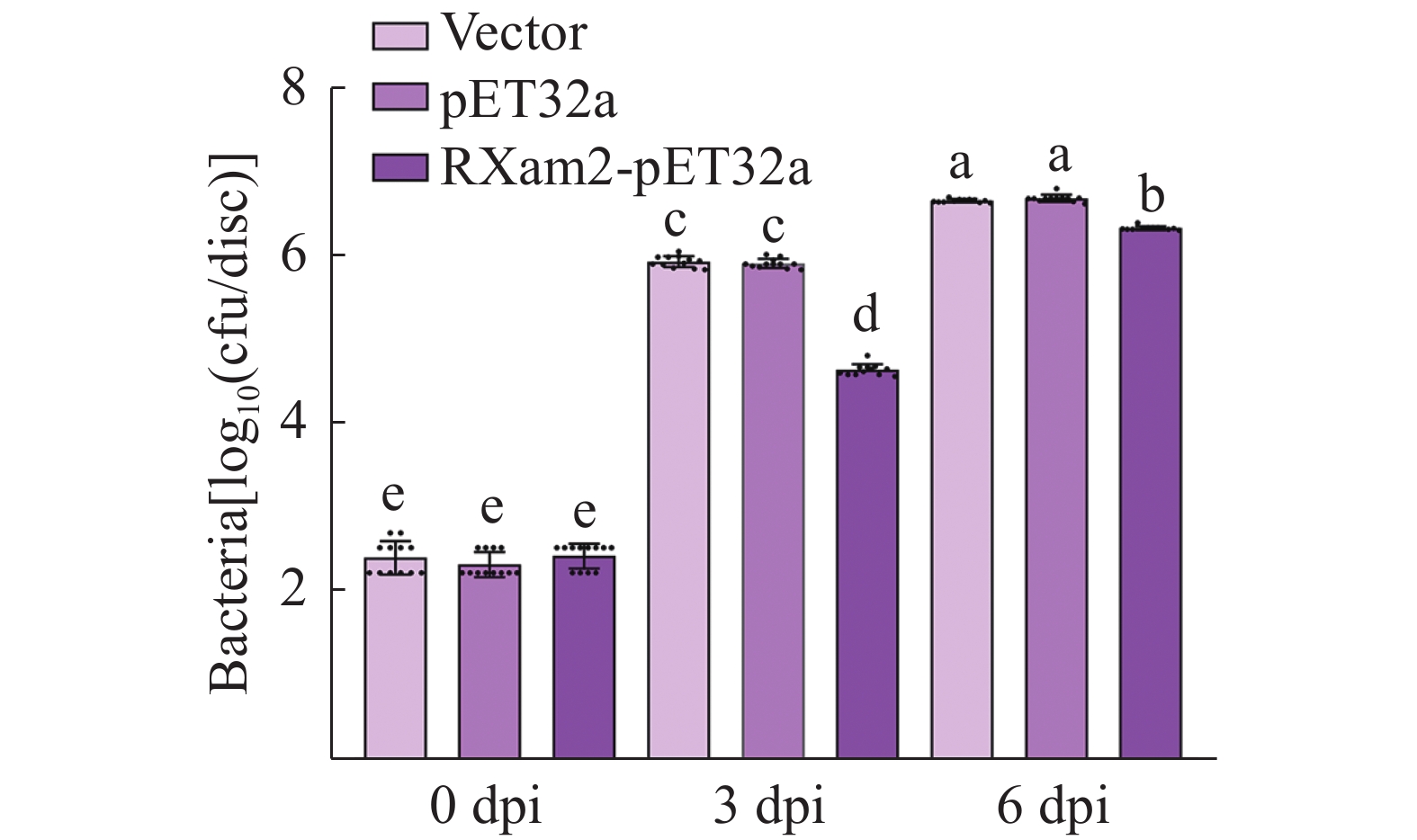
为探究MeRXam2蛋白对Xam繁殖的影响,本研究对木薯叶片接种了Xam病原菌,并对接种了Xam的木薯植株分别外源喷施水、pET32a蛋白和MeRXam2蛋白,检测病菌的繁殖情况,结果发现,对照组的细菌数显著高于实验组(图6),推测MeRXam2蛋白可能参与了木薯的抗病过程,并且外源喷施MeRXam2蛋白可有效减缓病原菌的繁殖。
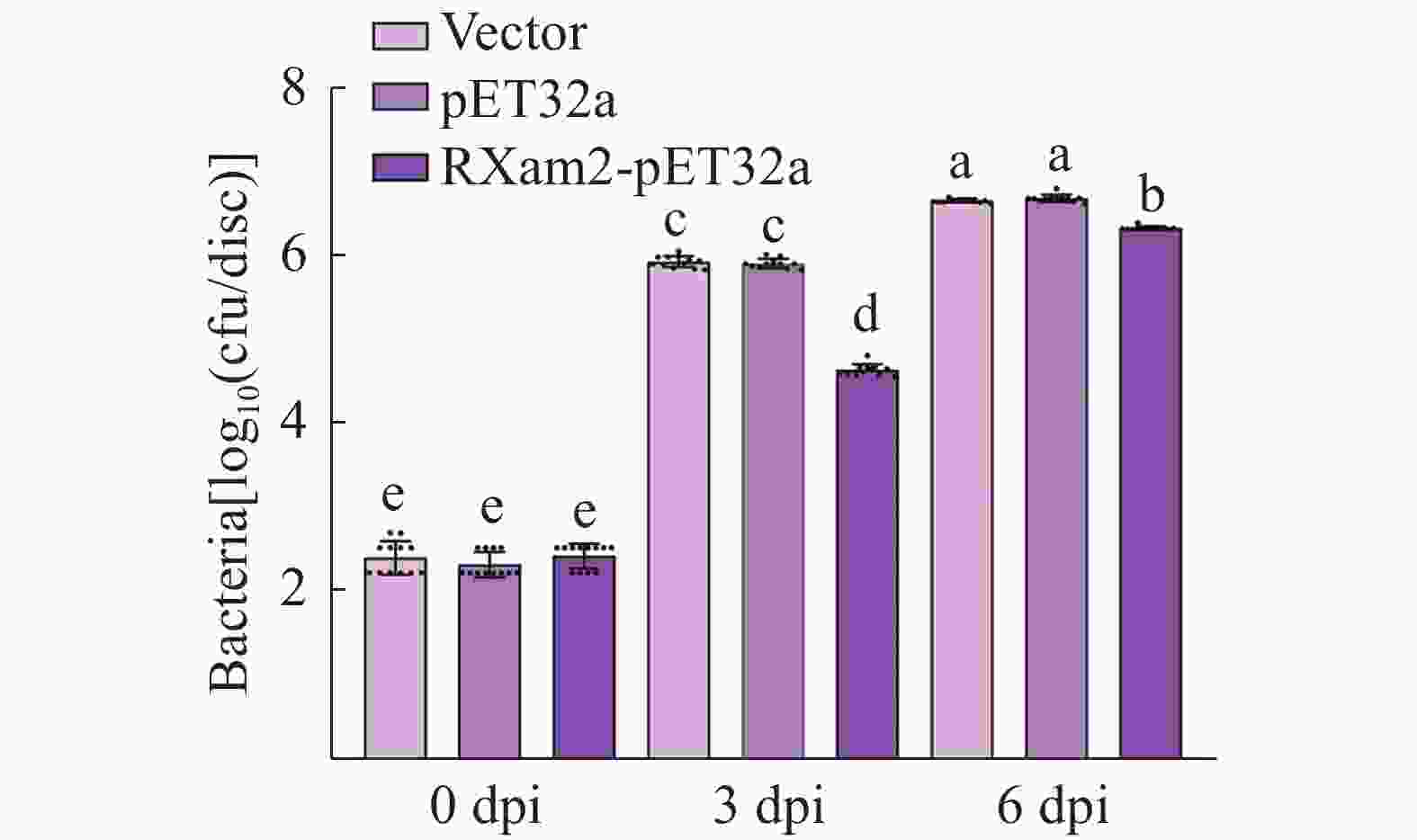
Figure 6. The influence of MeRXam2 protein on pathogenic bacteria
-
植物中编码NLR结构域的基因是植物中重要的抗病基因,能够识别病原菌微生物,调节免疫反应[27]。基于木薯细菌性枯萎病害的危害性,鉴定和分离木薯抗病基因具有重要意义[28]。因此挖掘出有利用价值的NLR基因,对提高木薯产量和保证粮食安全具重要意义[29]。
目前研究表明,不同物种中皆发现了具有NLR结构的蛋白,该结构蛋白常具有NBS和LRR结构域,如苜蓿(Medicago ruthenica)基因组中共鉴定出338个NLR基因[30],薯蓣(Dioscorea polystachya)基因组中鉴定出167个NBS-LRR基因[31],猕猴桃(Actinidia chinensis)基因组中鉴定了100个NBS-LRR基因[32],研究对12个蔷薇科基因组进行有关NBS-LRR基因搜索,筛选出2 188个NBS-LRR基因[33]。本研究中分析了木薯MeRXam2的蛋白结构,发现MeRXam2的C端含有一种典型的NLR结构-LRR结构域(520~
1186 aa),这表明MeRXam2是NLR基因。不同物种的NLR蛋白都与抗病性有关联,例如,水稻(Oryza sativa)[34]、拟南芥(Arabidopsis thaliana )[35]、小麦[36]等物种。在具有NBS-2的转化体的水稻中,其植株对叶瘟病能产生抗性,通过基因转化和标记辅助选择渗入易感籼稻品种中,赋予了籼稻株对叶片稻瘟病的抗性[34]。在拟南芥中ZAR1抗病小体,作为钙渗透性阳离子通道来触发免疫和细胞死亡[35]。在小麦中发现TaRACK1B能调节小麦花叶病毒感染的免疫反应,病毒可能会破坏植物NLR蛋白复合物,抑制NLR蛋白介导的广谱抗病毒免疫反应,从而导致小麦易受病毒感染[36]。与此一致的是,本研究发现MeRXam2可以参与到木薯抗病应答反应中。在有关蛋白与植物的抗病研究中,外源喷施超敏蛋白对葡萄霜霉病有抑制效果[37]。进一步对MeRXam2蛋白抗病功能分析中,Xam的繁殖受到外源喷施的MeRXam2蛋白的影响,MeRXam2蛋白可有效缓解细菌的侵染。NLR蛋白在体内同样也发挥了重要作用,在NLR蛋白在植物防御反应中,有通过充当守卫或诱饵,有作为下游信号转导成分,有调节蛋白结构等各种机制,进而影响植物的免疫反应[38]。后续可进一步研究MeRXam2在木薯体内的具体抗病分子机制。
综上所述,NLR是植物中重要的抗病基因家族,NLR蛋白对提高植物免疫力具有巨大潜在研究价值,本研究为木薯分子育种提供了理论依据和候选蛋白。
Analysis of prokaryotic protein expression and disease resistance function of cassava RXam2
DOI: 10.15886/j.cnki.rdswxb.20240147
- Received Date: 2024-09-13
- Accepted Date: 2024-12-31
- Rev Recd Date: 2024-11-30
- Available Online: 2025-01-22
- Publish Date: 2025-07-25
-
Key words:
- cassava /
- gene cloning /
- prokaryotic expression /
- immune response
Abstract: Cassava (Manihot esculenta Crantz) has important economic value. Exploring the key genes of cassava is of great significance for improving cassava quality. To mine disease-resistant genes of cassava, a specific immune receptor-related gene resistant to Xam (MeRXam2) was cloned, and its protein structure was analyzed. The analysis was found that the coding sequence (CDS) region of this gene is
| Citation: | TANG Luzhi, WU Jingyuan, CHEN Yuanlai, ZHU Binbin, ZHAO huiping. Analysis of prokaryotic protein expression and disease resistance function of cassava RXam2[J]. Journal of Tropical Biology, 2025, 16(4): 561-569. doi: 10.15886/j.cnki.rdswxb.20240147

|
 DownLoad:
DownLoad: