
-
斑翅果蝇(Drosophila suzukii Matsumura)别名铃木氏果蝇,隶属双翅目(Diptera),环列亚目(Cyclorrhapha),果蝇科(Drosophilidae)果蝇属 (Drosophila),是主要的水果果实害虫。斑翅果蝇传播范围广、速度快,自1916年在日本山梨县首次发现后迅速蔓延,已严重危害了美国、日本、俄罗斯等30多个国家的樱桃、蓝莓等水果产业,并被多个国家列为重要的检疫性害虫[1-3]。现如今我国大部分水果产区,广西、广东、云南、台湾等省均有危害记载,山东、辽宁、黑龙江等北方省份也相继报道过斑翅果蝇的危害情况[4-7]。斑翅果蝇寄主广泛,危害严重,危害对象包括樱桃、葡萄、柿子、杨梅、蓝莓等18科60多种水果[5, 8]。斑翅果蝇不同于其他果蝇,除取食落地果和受损果实外,其雌性成虫坚硬的锯齿状产卵器,能将成熟皮软的果实表皮刺破,在其内产卵,幼虫取食危害,造成严重的经济损失[5]。我国出口澳大利亚的油桃、葡萄等水果曾因为有可能携带斑翅果蝇而被要求实施检疫处理[9]。
由于斑翅果蝇具有传播广、繁殖快、危害严重等特点,目前对于防治斑翅果蝇最直接有效的手段还是化学防治[10]。而长期过量使用化学农药会导致昆虫对某些杀虫剂的敏感性明显降低而产生抗药性[11]。最新研究发现,在实验室药剂汰选下,斑翅果蝇可以在短时间内对氯氰菊酯、马拉硫磷和多杀菌素产生抗性[12-13]。昆虫产生抗药性的主要机制之一是其体内的代谢解毒酶的活性显著增高,将杀虫剂转化为极易溶于水的极性分子,通过一系列氧化还原作用、水解作用、轭合作用和基团转移作用,将其排出体外,从而增强对杀虫剂的解毒能力,这种机制称为代谢抗性[14]。参与代谢反应的解毒酶通常被分为5类:ABC转运蛋白(ATP-binding cassette transporter, ABCs)、细胞色素P450单加氧酶(cytochrome P450, CYPs)、谷胱甘肽-S-转移酶(glutathione S-transferases,GSTs)、羧酸酯酶(carboxylesterases, CarEs)、尿苷二磷酸葡萄糖醛酸转移酶(UDP-glucuronosyl transferases, UGTs)。昆虫的解毒代谢过程分为3个阶段:① 细胞色素P450单加氧酶(cytochrome P450, CYPs)参与第一阶段[15]。细胞色素P450酶系广泛分布于生物体内,主要由细胞色素P450、细胞色素b5、黄素蛋白-NADPH-P-450还原酶、黄素蛋白-NADH-细胞色素b5还原酶以及磷脂组成,是参与外源和内源化合物合成和分解的一种重要代谢酶系, CYPs 氧化代谢活性的增强是昆虫对拟除虫菊酯类杀虫剂产生抗性的重要机制[16] 。也有研究表明冈比亚按蚊对氯菊酯的抗性增强与其体内的CYP6P3基因表达过量有关[17]。② 3类解毒酶参与代谢解毒过程的第二阶段[18],谷胱甘肽-S-转移酶属于多功能的超家族解毒酶系,通常认为昆虫通过GSTs的共轭结合、脱氯化氢、螯合以及提高细胞的抗氧化活性等一系列作用来代谢外源有毒物质,从而产生抗药性[19]。羧酸酯酶作为一种重要的解毒酶,存在于昆虫头部、中肠、马氏管等组织中,它对外源生物分子的高亲和性和解毒作用是昆虫对有机磷类和氨基甲酸酯类杀虫剂产生抗性的重要原因[20]。尿苷二磷酸葡萄糖醛酸转移酶是一类广泛存在于动物、植物、细菌以及病毒内的多功能超家族酶系[21], 参与动物、植物、细菌和病毒等生物体内各种化合物的解毒作用,可以将核糖核苷酸中的糖基团催化形成亲水性化合物并有效排出体外[22] 。③ 第三阶段由ABC转运蛋白(ATP-binding cassette transporter, ABCs)参与代谢解毒过程。ABC转运蛋白能够直接将极性化合物或复合物以及其他外源有毒物质泵出细胞外,而不需要经过其他酶的修饰,从而起到排毒和保护昆虫的作用[23]。家蚕体内ABC转运蛋白的表达上调可以降低其对杀虫剂的敏感性[24]。
通过分析基因进化关系以及蛋白保守结构域可以鉴定基因的类别、解析基因的功能,随着全基因组测序技术的不断进步和发展,以及大量昆虫全基因组测序的完成,此类研究日益增多。梅洋、Yang等分别通过草地贪夜蛾sf9细胞系的基因组数据和脑转录组数据,鉴定并注释了草地贪夜蛾的解毒酶相关基因家族[25-26];基于基因组和转录组数据,尹传林等解析了二化螟的抗药性相关基因以及寄生蜂细胞色素P450的基因家族[27- 28]。同时此类研究也被应用于昆虫化学感受相关基因和表皮蛋白基因等方面[29,-30]。但在斑翅果蝇中还未有对其解毒代谢相关基因的鉴定与分析,本研究拟从NCBI中获得斑翅果蝇的转录组数据,对其与代谢解毒相关的5个主要基因家族进行分类,并分析它们的保守结构域与氨基酸残基,以期为斑翅果蝇的防治提供新的思路和途径。
-
使用TBtools v1.085软件从NCBI 数据库(
https://www.ncbi.nlm.nih.gov/nuccore/GCA_000472105.1/GCF_000472105.1 )下载斑翅果蝇的转录组及参考转录组序列。同时下载获得其他29种果蝇的UGTs核苷酸序列和21种果蝇的CESs核苷酸序列。根据黑腹果蝇的基因组注释方法和结果将斑翅果蝇的解毒基因归类为CYPs、CarEs、GSTs、ABCs和UGTs五个家族[31]。在斑翅果蝇转录组数据中搜索得到CYPs、CarEs、GSTs、ABCs和UGTs的核苷酸序列,通过NCBI核苷酸数据库中的同源性blast功能进行人工验证,将筛选标准设定为e值≤ 1e−5,识别百分比≥ 50%[32]。采用 NCBI Open Reading Frame Finder 预测5种解毒代谢相关基因的开放阅读框(ORF)(https://www.ncbi.nlm.nih.gov/orffinder/ ),选出具有完整开放阅读框的基因,使用EditSeq软件将已鉴定的基因翻译成氨基酸序列。 -
将最终确定的 CYPs、ABCs、GSTs、CarEs、UGTs 氨基酸序列通过MEGA7. 0 软件中的ClustW进行多序列比对[33],将比对后的文件转化为meg格式,基于最大似然法,本研究使用Mega-X软件构建具有1 000个bootstrap值的系统进化树,并通过iTOL工具(
https://itol.embl.de/itol.cgi )进一步润色和美化(CarEs和UGTs由于数据太少,将其与其他果蝇的同源基因进行比对)。 -
将具有完整开放阅读框的基因使用模块化结构研究工具SMART(
http://smart.embl.de/ )用于预测5种解毒代谢基因的保守结构域[34],并将CYPs、ABCs、GSTs的氨基酸序列分别上传至在线工具MEME(https://meme-suite.org/meme/ ),分析此3种斑翅果蝇解毒代谢基因中保守的氨基酸残基[35]。 -
经过序列比对分析,从斑翅果蝇转录组数据中鉴定出58条具有完整开放阅读框的P450s序列,平均长度为1 847 bp,编码428~595个氨基酸;28条编码GSTs的核苷酸序列,平均长度为1 005 bp,编码209~557个氨基酸;26条编码ABCs 的核苷酸序列,平均长度为3034 bp,编码604~2201个氨基酸。从NCBI中下载的29种果蝇的具有完整开放阅读框的UGTs核苷酸序列平均长度为1 578 bp,编码497~540个氨基酸;21种果蝇的CarEs核苷酸序列平均长度为1 999 bp,编码602~681个氨基酸。
-
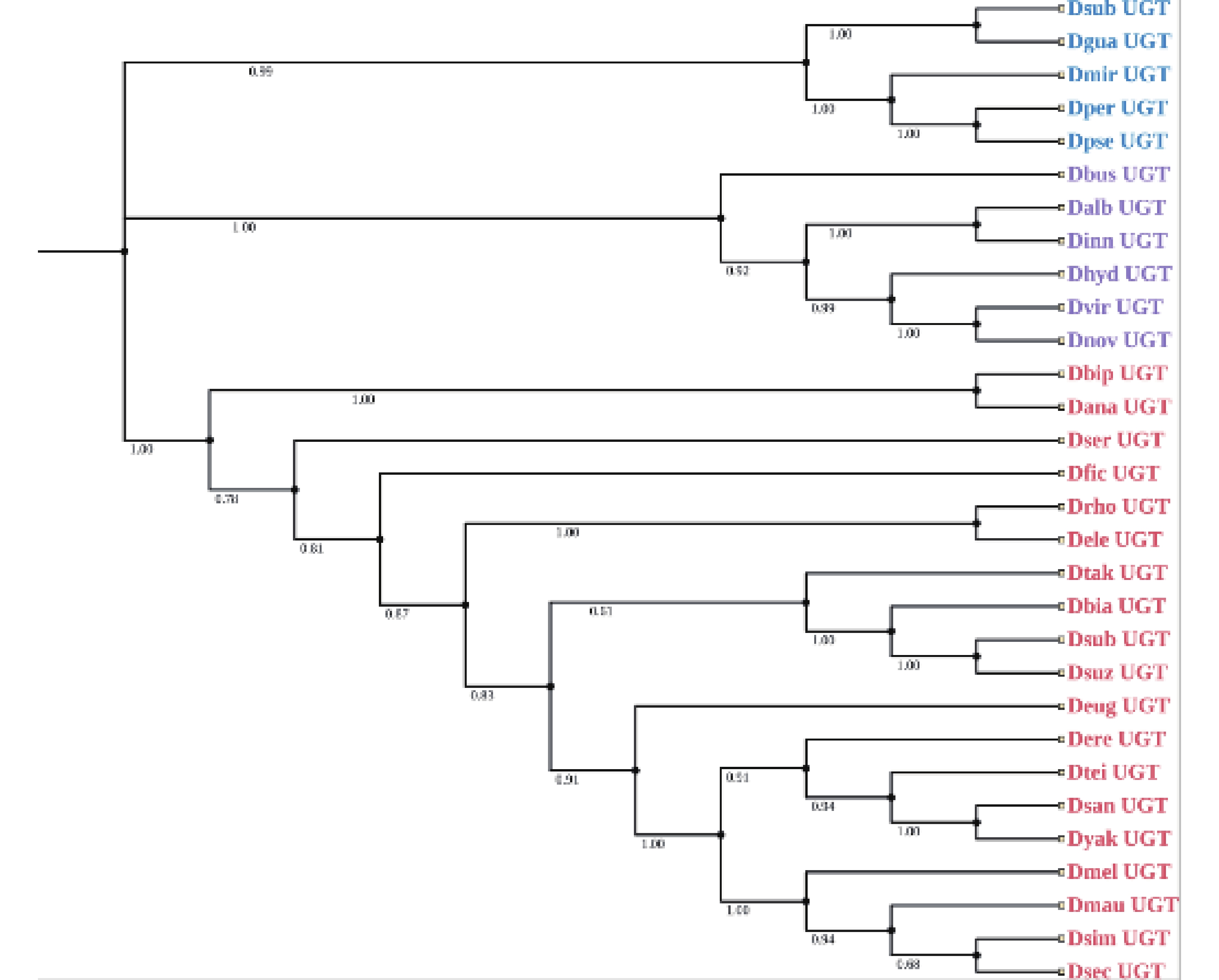
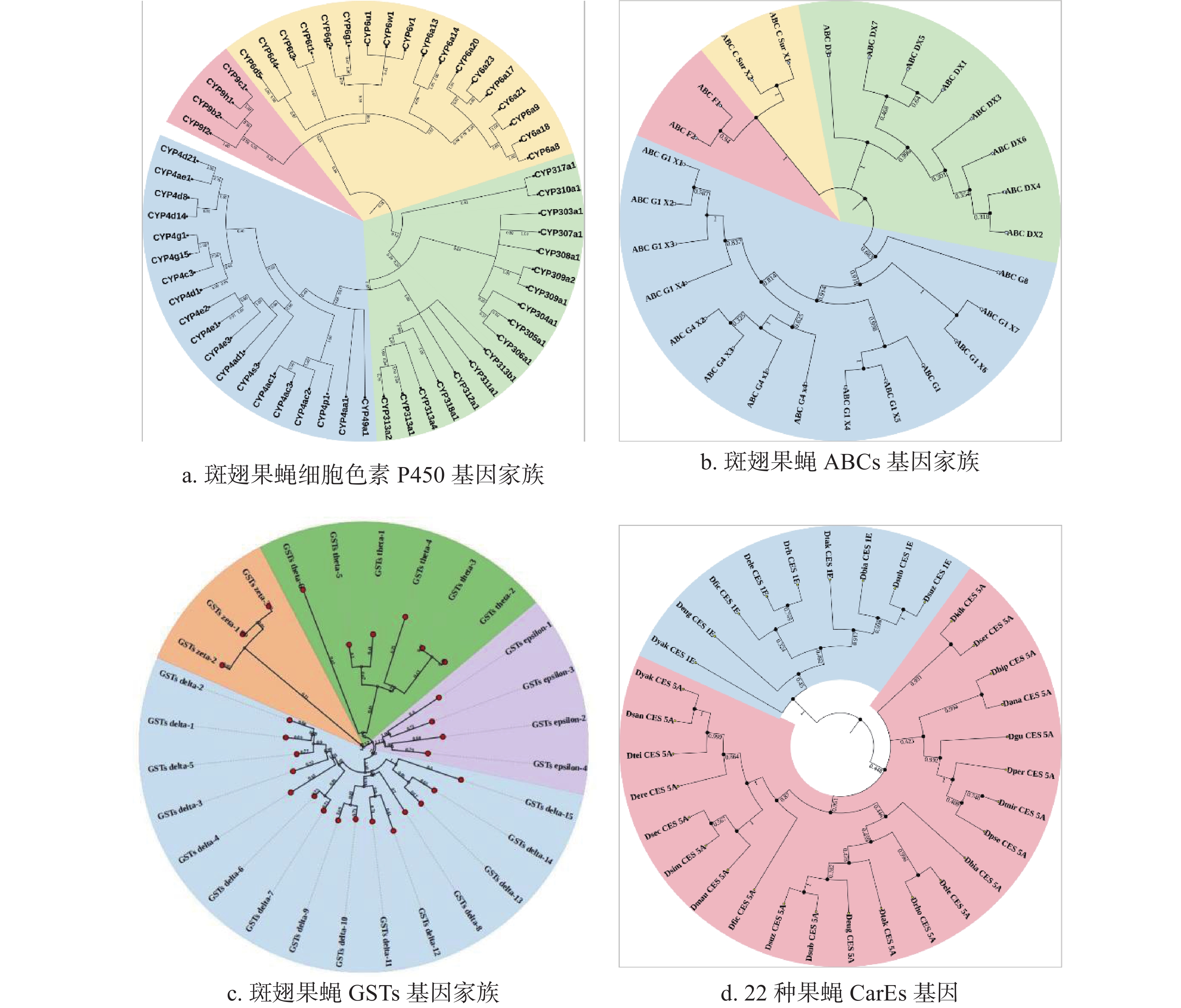
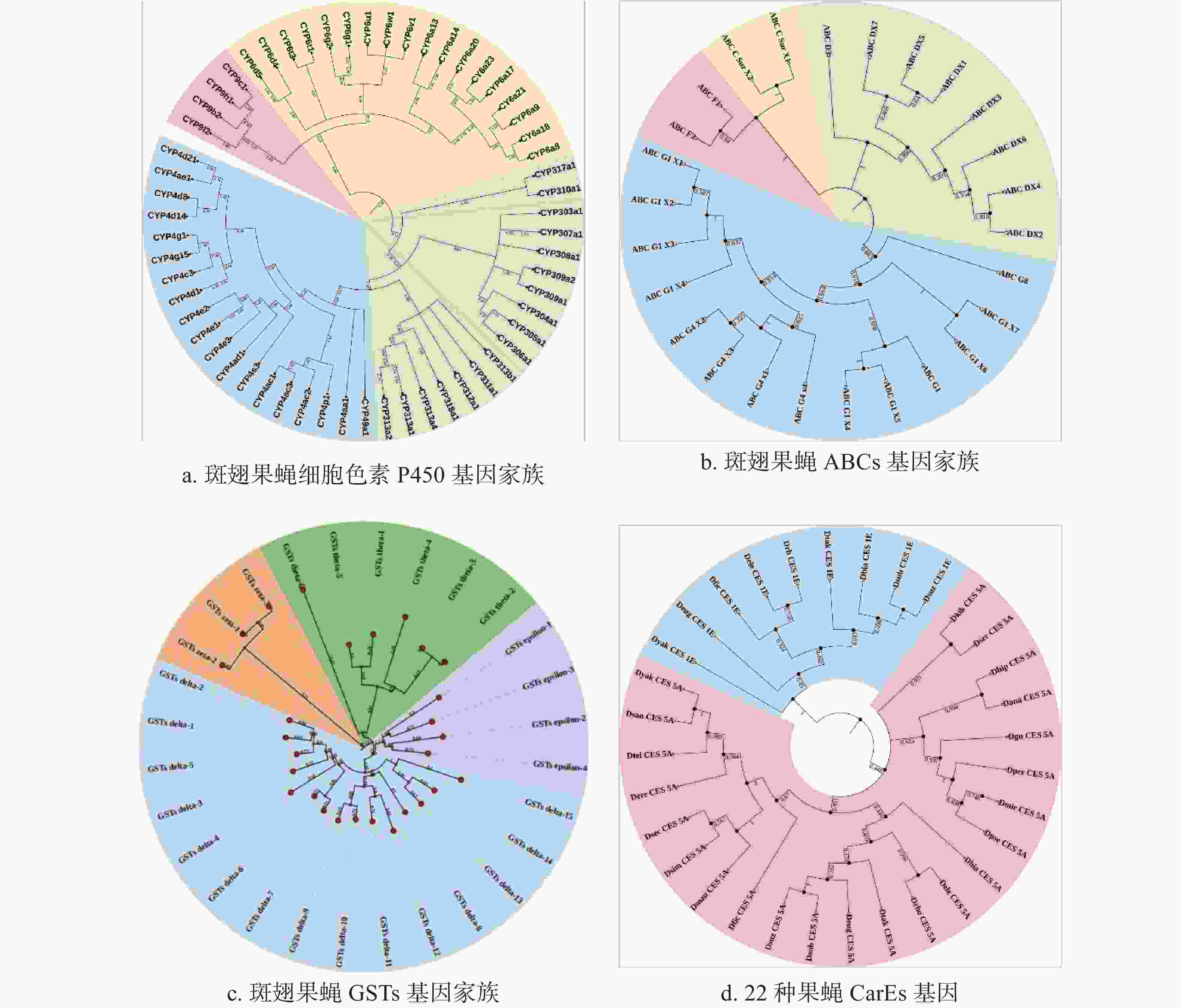
基于CYPs、GSTs、ABCs的保守区域,本研究利用这些氨基酸序列分别构建了系统发育树(图1-a、图1-b、图1-c)。结果显示58个CYPs基因分布于CYP3、CYP4、CYP6、CYP9四个进化分支,其中包含19条序列的CYP4家族构成了发育树的最大分支,而CYP9分支最小,只有4条序列;而ABCs基因系统发育树由4个分支组成,其中ABC-G构成的分支最为庞大,包含14条序列;28个GSTs基因分布于GST-Delta、GST-Epsilon、GST-Theta以及GST-Omega四个进化分支,分别为其中最大的进化分支为GST-Delta。由于数据库中的CarEs和UGTs基因太少,本研究用斑翅果蝇的CES基因与其他21种果蝇的CESs氨基酸序列构建系统发育树,结果显示果蝇的CarEs基因分为CES-1E和CES-5A两个进化枝,其中斑翅果蝇与亚艳丽果蝇(Drosophila subpulchrella)同源性最高(图1-d)。同时,本研究将斑翅果蝇的UGTs基因与其他29种果蝇的UGTs氨基酸序列构建系统发育树,结果显示30种果蝇的UGTs基因被分为3个进化枝,在同一个进化枝中斑翅果蝇和亚艳丽果蝇(D. subpulchrella)的遗传距离最近(图2)。
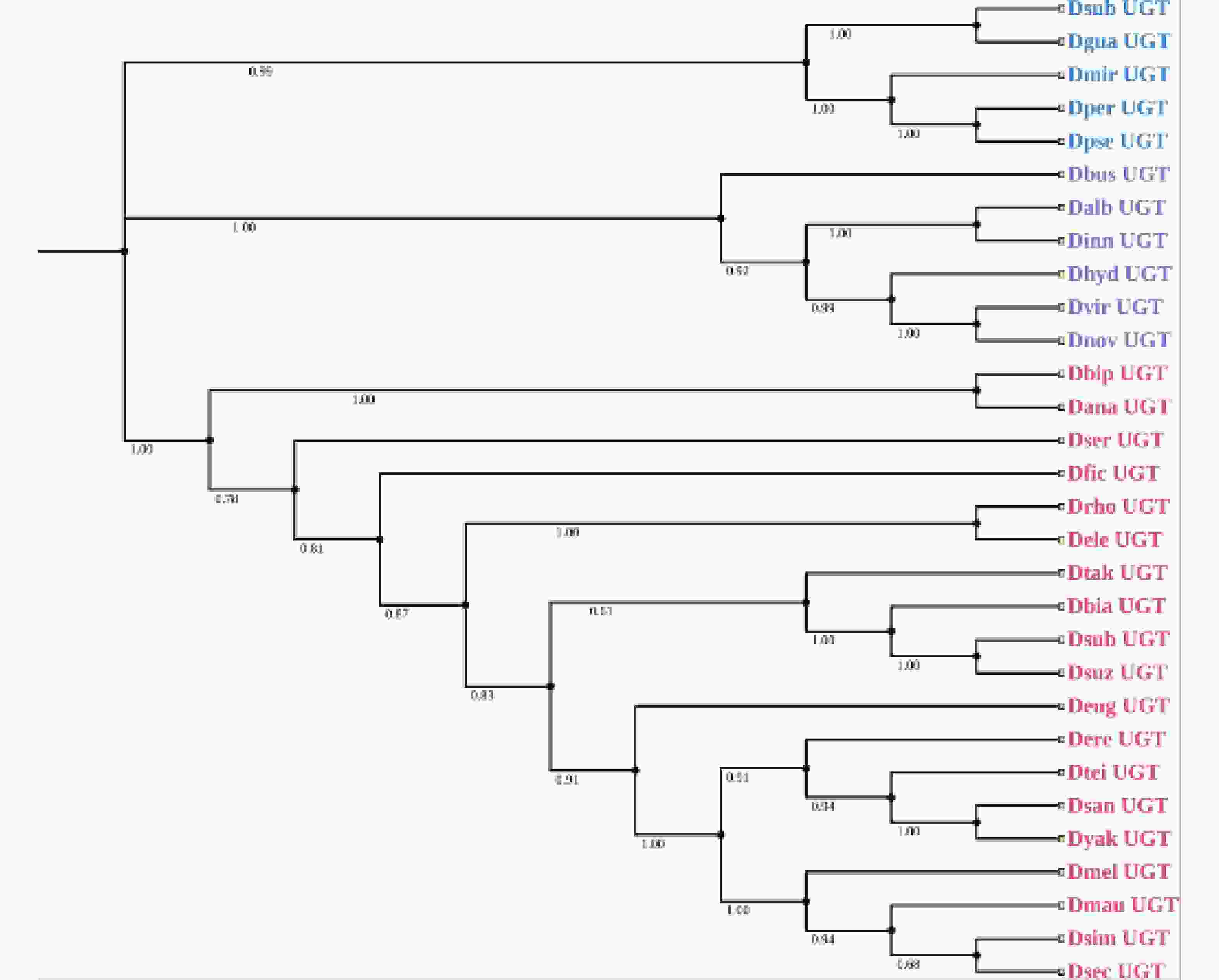
-
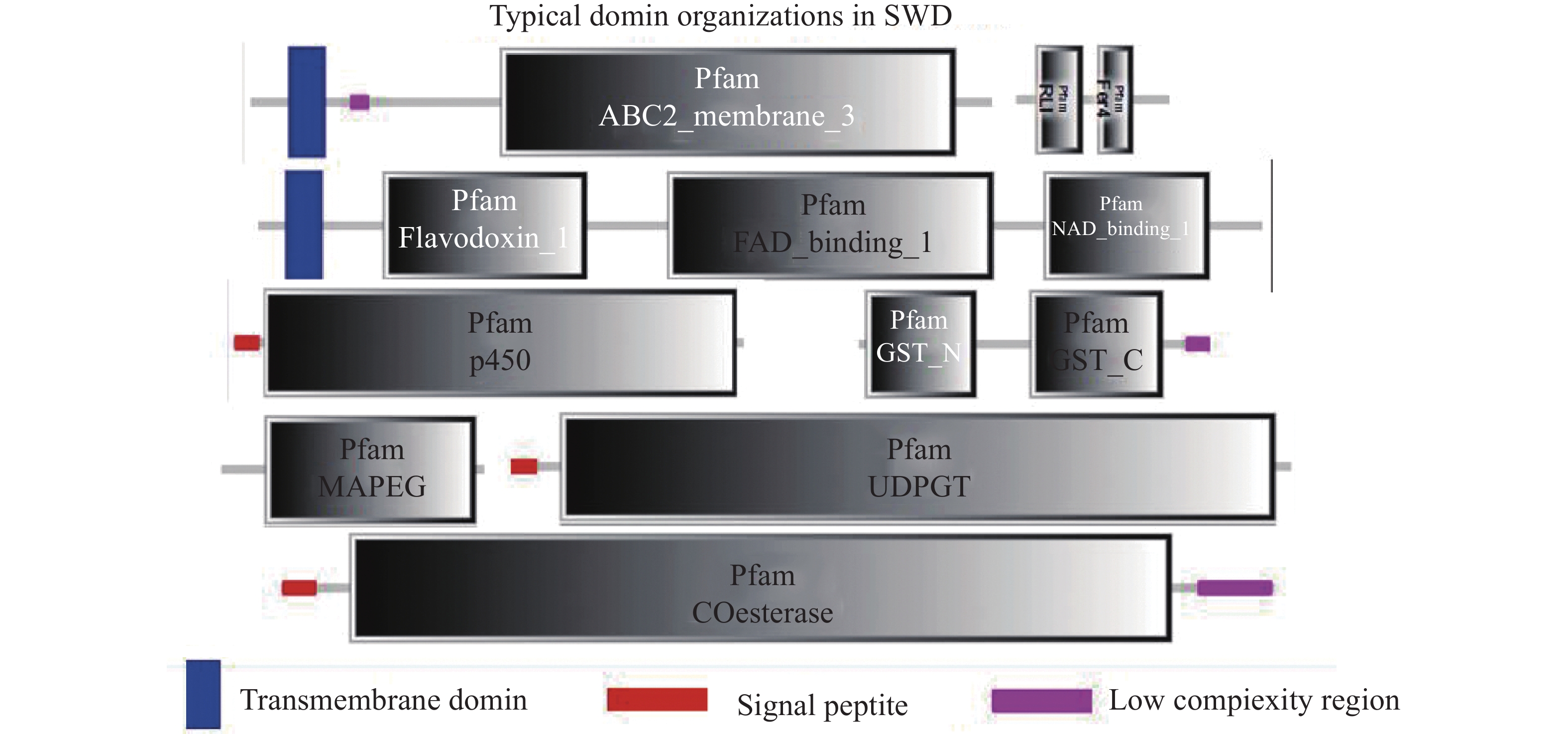
通过SMART工具,本研究分析了斑翅果蝇的CYPs、CarEs、GSTs、ABCs和UGTs五种解毒酶基因的蛋白结构域。分析结果显示在CYPs中除了信号肽(signal peptide)和细胞色素P450蛋白的保守区域以外,还包含FAD结合域(FAD-binding)、NAD结合域(NAD-binding)、以及起电子传递作用的黄素氧还蛋白(flavodoxin)。在ABCs中除了包含ABC转运蛋白保守区域的ATP结合盒(ABC-membrane)外还包括核糖核酸内切酶RNase L抑制蛋白N-末端的RLI结构域以及在多种代谢反应中介导电子转移的4Fe-4S的铁氧化还原蛋白(Fer4)。在GSTs中包括谷胱甘肽-S-转移酶的N-末端结构域、C-末端结构域以及类花生酸和谷胱甘肽代谢中的膜相关蛋白MAPEG(Membrane Associated Proteins in Eicosanoid and Glutathione metabolism)。在CarEs和UGTs中只发现了信号肽和它们各自的保守区域:羧酸酯酶-结构域(Coesterases)以及UDP糖基转移酶蛋白(UDPGT)(图3)。此外,根据在线工具MEME的分析结果本研究分别在斑翅果蝇的CYPs中鉴定出几个高度保守的氨基酸残基EEGGKKRNDFLDLLJZLKKEG(图4-a);在ABCs中鉴定出的保守氨基酸残基为DCPSASNPADYII(图4-b);在GSTs中鉴定出的保守氨基酸残基为LYPKDLVKRAVVDQRLHFE(图4-c)。
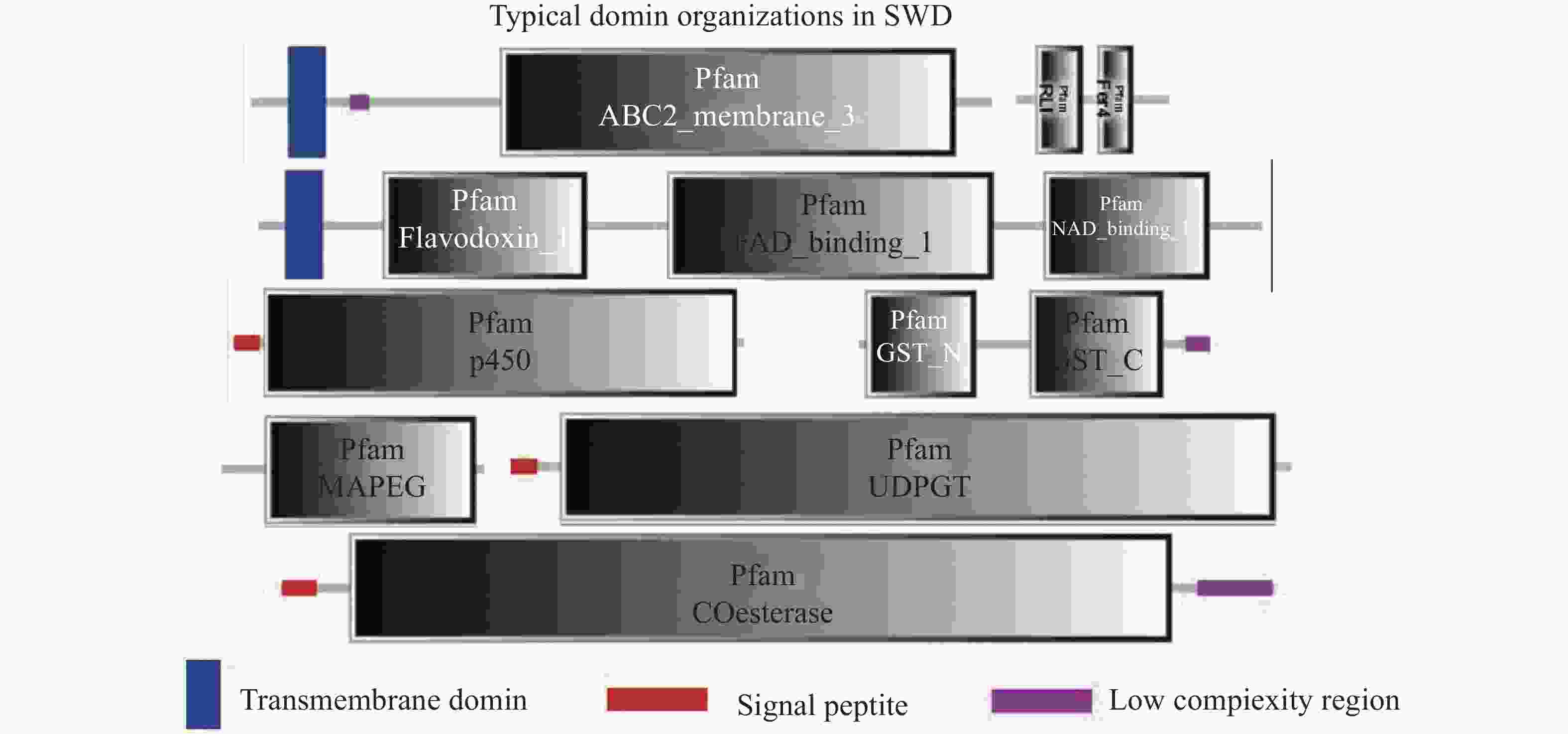

-
代谢抗性是近几十年来被广泛研究的昆虫产生抗药性的主要机理之一[36]。随着功能基因组技术在昆虫抗药性研究中的应用,一些与解毒代谢相关的基因在昆虫中被鉴定出来,一般情况下,解毒酶基因包括CYPs、ABCs、CarEs、GSTs、UGTs等[37]。研究结果表明昆虫的代谢解毒能力与其寄主范围以及食性有重要关系[38]。细胞色素P450作为昆虫体内最大的解毒酶系之一 ,已被证明是大多数农业害虫对杀虫剂产生高抗药性以及交互抗性的主要原因[39]。在家蚕中鉴定出82 个细胞色素P450基因,而在食性更杂的草地贪夜蛾中则鉴定出213个细胞色素P450 基因[40]。本研究在斑翅果蝇基因组数据中共鉴定出58个包含完整开放阅读框的细胞色素P450 基因,其中CYP6家族包含18个基因,而CYP6家族被认为与昆虫抗药性关系最为密切[39]。已有研究报道在冈比亚按蚊抗氯菊酯品系中P450s基因CYP6Z1的表达量是敏感品系的11倍;在黑腹果蝇抗DDT品系中发现 CYP6G2基因高表达, 其表达量是敏感品系的10 倍~ 100倍[41]。在本研究中同样发现了CYP6G2基因,因此本研究结果将为探索斑翅果蝇CYPs解毒的分子机制提供重要参考。此外研究发现的几个细胞色素P450基因的保守氨基酸位点可能对其功能起重要作用。抗性昆虫主要通过提高羧酸酯酶的表达量和编码氨基酸发生突变引起结构的变化,导致酶促作用增强两种机制来提高其对杀虫剂的抗药性[42 -43]。酯酶基因表达量增强导致抗性产生的机制,最早在桃蚜(Myzus persicae)中发现,在抗性桃芽品系中酯酶活性明显高于敏感品系[44]。酯酶氨基酸突变导致的抗性最早在家蝇(Muscadomestica)对有机磷的抗药性研究中发现,在6个抗有机磷家蝇品系中均有一个突变酯酶E3G137D[45]。而UDP-葡萄糖醛酸转移酶通过调节UGTs的表达量改变昆虫对杀虫剂的抗药性[46]。研究表明,当UGTs基因表达量提高时,马铃薯叶甲对吡虫啉的抗性会随之增强;而当褐飞虱的UGT-1-7和UGT2B10基因被抑制后,其对吡虫啉的敏感性会增强[47- 48]。谷胱甘肽 S-转移酶同样是一个多功能的超家族解毒酶系,可以使昆虫对所有主要类别的杀虫剂产生抗性。第一个昆虫的GSTs基因是从黑腹果蝇中克隆而出[49],在黑腹果蝇基因组中共发现37 个GSTs基因[19]。马铃薯甲虫、柑橘全爪螨、禾谷缢管蚜等昆虫中的GSTs基因与抗药性的关系也被研究报道[50-52]。本研究从斑翅果蝇转录组中发现28个GSTs基因,其功能是否与抗药性有关还有待研究。
经典的昆虫代谢抗性研究主要集中在细胞色素 P450 、羧酸酯酶以及谷胱甘肽S-转移酶3个家族[18]。而随着越来越多的昆虫全基因组测序的完成,有关 ABC 转运蛋白介导的代谢抗性的研究也日益增多。研究发现在小菜蛾(Plutella xylostella)中有100个ABC 转运蛋白基因,与敏感品系相比,ABCA,ABCC,ABCG,ABCH和ABCF家族的成员在小菜蛾抗毒死蜱品系中相对高表达[53];在黑腹果蝇中发现了56个ABC 转运蛋白基因,其中,ABCB、ABCC和ABCG基因家族主参与外源有毒物质的代谢,与杀虫剂抗性有关[54]。本研究在斑翅果蝇中发现了26个ABC 转运蛋白基因,包括ABCC、ABCD、ABCF、ABCG四个亚家族,其中ABCG亚家族最为庞大,这一结果将为斑翅果蝇ABCs的解毒机制提供重要参考。同时在ABC 转运蛋白中发现的几个保守氨基酸残基将为开发新农药提供潜在的靶标位点。
Identification and analysis of detoxification genes in Drosophila suzukii Matsumura
DOI: 10.15886/j.cnki.rdswxb.20230009
- Received Date: 2023-02-04
- Accepted Date: 2023-03-16
- Rev Recd Date: 2023-03-10
- Available Online: 2023-10-28
- Publish Date: 2023-11-24
-
Key words:
- Drosophila suzukii /
- metabolic detoxification gene /
- phylogenetic /
- protein domain /
- conserved motif
Abstract: Drosophila suzukii Matsumura is a worldwide fruit pest. Based on the transcriptome of D. suzukii, five metabolic detoxification genes were identified by using sequence analysis, phylogenetic analysis, structural domain analysis and conserved motif analyses. These five genes include ATP-binding cassette transporter (ABCs), cytochrome P450 (CYPs), glutathione S-transferases (GSTs), carboxylesterases (CarEs) and UDP-glucuronosyl-transferases (UGTs). Phylogenetic analysis showed that CYPs genes were classified into four clans, CYP3, CYP4, CYP6 and CYP9, while ABCs genes fell into four clans, ABCC, ABCD, ABCF and ABCG. GSTs genes were classified into GST-Delta, GST-Epsilon, GST-Theta and GST-Zeta. The CarEs genes and UGTs genes of D. suzukii were the closest to those of Drosophila subpulchrella. Structural domain and conserved motif analyses showed that CYPs had a conserved P450 domain; ABCs had a conserved ABC2 domain; GSTs had a conserved GST domain; CarEs had a conserved Coestrases domain; UGTs had a conserved UDPGT domain. Conservative motifs of CYPs were EEGGKKRNDFLDLLJZLKKEG. Conservative motifs of ABCs were DCPSASNPADYIIE. Conservative motifs of GSTs were LYPKDLVKRAVVDQRLHFE.
| Citation: | WANG Zhenglei, LIU Kaiyang, YUAN Linlin, LI Fen, WU Shaoying. Identification and analysis of detoxification genes in Drosophila suzukii Matsumura[J]. Journal of Tropical Biology, 2023, 14(6): 660-667. doi: 10.15886/j.cnki.rdswxb.20230009

|
 DownLoad:
DownLoad: