-
斑节对虾(Penaeus monodon)属甲壳纲(Crustacea)十足目(Decapoda)对虾科(Penaeidae)对虾属(Penaeus),又名黑虎虾、草虾,是当今世界三大经济对虾养殖品种之一。斑节对虾是对虾属中体型最大的种类,分布广泛,主要在印度洋以及太平洋区域[1]。自20世纪80年代起,斑节对虾养殖规模逐渐扩大。近年来,养殖环境污染引起的多种细菌病、病毒病等疾病大规模爆发,严重制约了斑节对虾养殖产业的健康发展。其中,细菌病主要是由副溶血弧菌(Vibrio parahaemolyticus)、哈维弧菌(V. harveyi)等十几种病原菌引发[2]。有研究显示,细菌脂多糖 (LPS) 是几乎所有革兰氏阴性细菌外表面膜的主要成分,并且是多种真核生物先天免疫反应的极强刺激剂[3]。对虾的先天性免疫是维持虾健康的主要屏障,深入研究斑节对虾先天免疫防御对控制对虾疾病爆发和健康养殖具有重要意义。ML蛋白 (MD-2-related lipid-recognition, MD-2脂质识别蛋白) 家族是一类广泛分布于动植物和真菌等生物体内的多成员的蛋白家族,MD-2 对于细菌脂多糖 (LPS) 介导的信号传导过程和后续的促炎因子分泌过程至关重要[4]。这些蛋白分子较小,通常都具有1个约150个氨基酸残基的ML结构域,其中,至少含有2对保守的半胱氨酸[5]。不同的ML蛋白具有不同的氨基酸序列,从而形成具有差异的中央疏水腔,可结合不同的脂类配体,产生脂类识别的特异性,进一步调节不同的生物学功能[6]。ML蛋白家族成员众多,因脂质识别特异性导致其在不同生物体的分布位置及行使功能有很大不同[7]。对脊椎动物中ML的研究较多,发现其在先天免疫和脂类代谢上发挥重要作用,例如人类MD-2参与Toll样受体4 (Toll-like receptors 4, TLR4)识别细菌脂多糖LPS,还可以直接结合LPS[8],而人类C型尼曼匹克蛋白2 (Niemann-Pick type C2 protein, NPC2)具有很强的胆固醇结合能力,能快速转运胆固醇至膜受体从而调节细胞内的脂质平衡[9]。无脊椎动物中,ML的功能还未分化和明确,可能行使更为复杂多样的生物学功用,研究发现,黑腹果蝇(Drosophila melanogaster)的NPC2a就同时具备了相当于脊椎动物的MD-2和NPC2两者的功能[10]。此外,ML家族还有相当多功能不明确的蛋白有待深入研究。近年来,对无脊椎动物ML基因的研究不断深入,发现ML蛋白参与了宿主和病毒的相互作用[11]。在甲壳动物中,凡纳滨对虾(Litopenaeus vannamei)的LvML蛋白可能参与抗菌和抗病毒的先天免疫过程[12];日本囊对虾(Marsupenaeus japonicus)的ML不仅通过识别WSSV的脂质成分参与先天免疫应答,还可能与幼体发育中的变态及蜕皮有一定的关联[13-14]。本研究基于斑节对虾的2个ML基因的相似片段,克隆获得了它们的cDNA全序列,分析了其序列特征,并利用实时荧光定量PCR分析目的基因蛋白在各组织的表达分布情况,为进一步了解ML家族基因在斑节对虾先天免疫中的作用提供资料。
-
供试的斑节对虾(Penaeus monodon)来自海南省海洋与渔业科学院琼海科研基地高位池(20 m×20 m×2 m)的养殖群体,平均单只体长为(13.24±0.35)cm,平均单只的体质量为(19.45±0.61)g,暂养于水泥池(9.7 m×1.5 m×1.3 m)中。暂养7 d后,随机选取6尾身体完整无损伤的健康雌性个体,取其血细胞、眼柄、心、鳃、胃、肠、肝胰腺和肌肉,迅速置于RNAlater (ThermoFisher ,USA)中4 ℃过夜后,−20 ℃长期保存备用。
RNeasy Mini Kit购自德国Qiagen。2×Taq PCR StarMix with Loading Dye、2×SuperStar Plus Mix with Loading Dye、StarPrep Gel Extraction Kit和EZ-Blunt Zero pTOPO II Cloning Kit等试剂盒均购自北京康润诚业生物科技有限公司(GenStar)。SMARTer RACE 5′/3′ Kit、PrimerScript™ RT reagent Kit with gDNA Eraser(Perfect Real Time)、SYBR® Premix Dimer Eraser™(Perfect Real Time)均购自宝生物工程(大连)有限公司。
-
以斑节对虾2个ML基因相似片段,利用引物设计软件Primer Premier 5.0和分析软件Oligo 6.0设计引物进行cDNA全序列的克隆[15]。以斑节对虾延伸因子1α基因 (EF-1α)作为内参基因,基于获得的PmML1与PmML2的全长cDNA序列设计qRT-PCR引物。所有引物(表1)由深圳华大基因有限公司合成。
引物名称 引物序列(5′−3′) 用途 PmML1F1 AGCCCACAATCATGCGACAG PmML1 cDNA扩增 PmML1R1 GAGGTTCCTGAACCTGGCTATAC PmML1 cDNA扩增 PmML2F1 GTGTAGTATACCATGGCTGCCTC PmML2 cDNA扩增 PmML2R1 TGAGTCTGCGTTGTCCTTACAAG PmML2 cDNA扩增 PmML1FF1 TTGACTCGCATGTGAGGTGG PmML1 3′-RACE PmML1RR1 GGGTGACTTCCCCTGCTATG PmML1 5′-RACE PmML2FF1 CTCTGTATGGGCACGAGAGC PmML2 3′-RACE PmML2RR1 CAGGATATCGCCGCCTCATC PmML2 5′-RACE QPmML1F1 TTGACTCGCATGTGAGGTGG qRT-PCR目的基因扩增 QPmML1R1 GGGTGACTTCCCCTGCTATG qRT-PCR目的基因扩增 QPmML2F1 CTCTGTATGGGCACGAGAGC qRT-PCR目的基因扩增 QPmML2R1 CAGGATATCGCCGCCTCATC qRT-PCR目的基因扩增 EF-1αF1 AAGCCAGGTATGGTTGTCAACTTT qRT-PCR内参基因扩增 EF-1αR1 CGTGGTGCATCTCCACAGACT qRT-PCR内参基因扩增 -
使用RNeasy Mini Kit提取斑节对虾肝胰腺组织总RNA,取5 μL RNA进行1%琼脂糖凝胶电泳检测完整性,并使用ND-1000超微量紫外分光光度计检测RNA质量。使用StarScript II First-strand cDNA Synthesis Mix With gDNA Remover反转录制备cDNA模板。
分别利用1对引物扩增和验证斑节对虾PmML1、PmML2的序列片段,PCR反应体系(50 μL):无菌双蒸水(DDW) 22 μL,12×Taq PCR StarMix 25 μL,正反向引物分别1 μL,肝胰腺cDNA模板1 μL。PCR反应程序:95 ℃ 5 min;95 ℃ 30 s, 60 ℃ 30 s,72 ℃ 2 min,35个循环;72 ℃ 5 min。PCR产物使用StarPrep Gel Extraction Kit回收目的条带,使用EZ-Blunt Zero pTOPO II Cloning Kit连接,连接产物加入到50 μL Trans5α Phage Resistant Chemically Competent Cell中转化,菌液涂于氨苄青霉素(Amp+)抗性平板上, 37 ℃培养过夜,随机挑取单菌落M13引物进行菌液PCR鉴定,挑取阳性克隆送测序获得目的片段序列。基于验证的PmML1、PmML2的序列片段设计的RACE巢式引物,根据SMARTer RACE 5′/3′ Kit 试剂盒的方法说明进行RACE实验的cDNA模板制备及PmML1、PmML2基因的3′末端序列和5′末端序列的扩增测序。
-
斑节对虾ML基因的ORF区域及氨基酸序列使用NCBI的ORF Finder在线程序(
https://www.ncbi.nlm.nih.gov/orffinder/ )确定[16]。利用PSORT II Prediction(https://psort.hgc.jp/form2.html )进行蛋白质亚细胞定位预测[17]。使用MAFFT在线软件(https://mafft.cbrc.jp/alignment/server/ )进行氨基酸序列的多重比对分析[18]。序列的比对、理化性质和系统发生树的构建等参照文献[19]的方法,蛋白的二级结构、功能结构域和糖基化位点的预测等参照文献[20]的方法。 -
提取斑节对虾血细胞、眼柄、鳃、心、胃、肠、肝胰腺与肌肉等8种组织总RNA,制备cDNA模板及实时荧光定量PCR检测各组织表达分布的方法参照文献[17]的方法进行,表达数据采用2−ΔΔCt法[21],用Excel2019软件分析和展示检测结果。
-
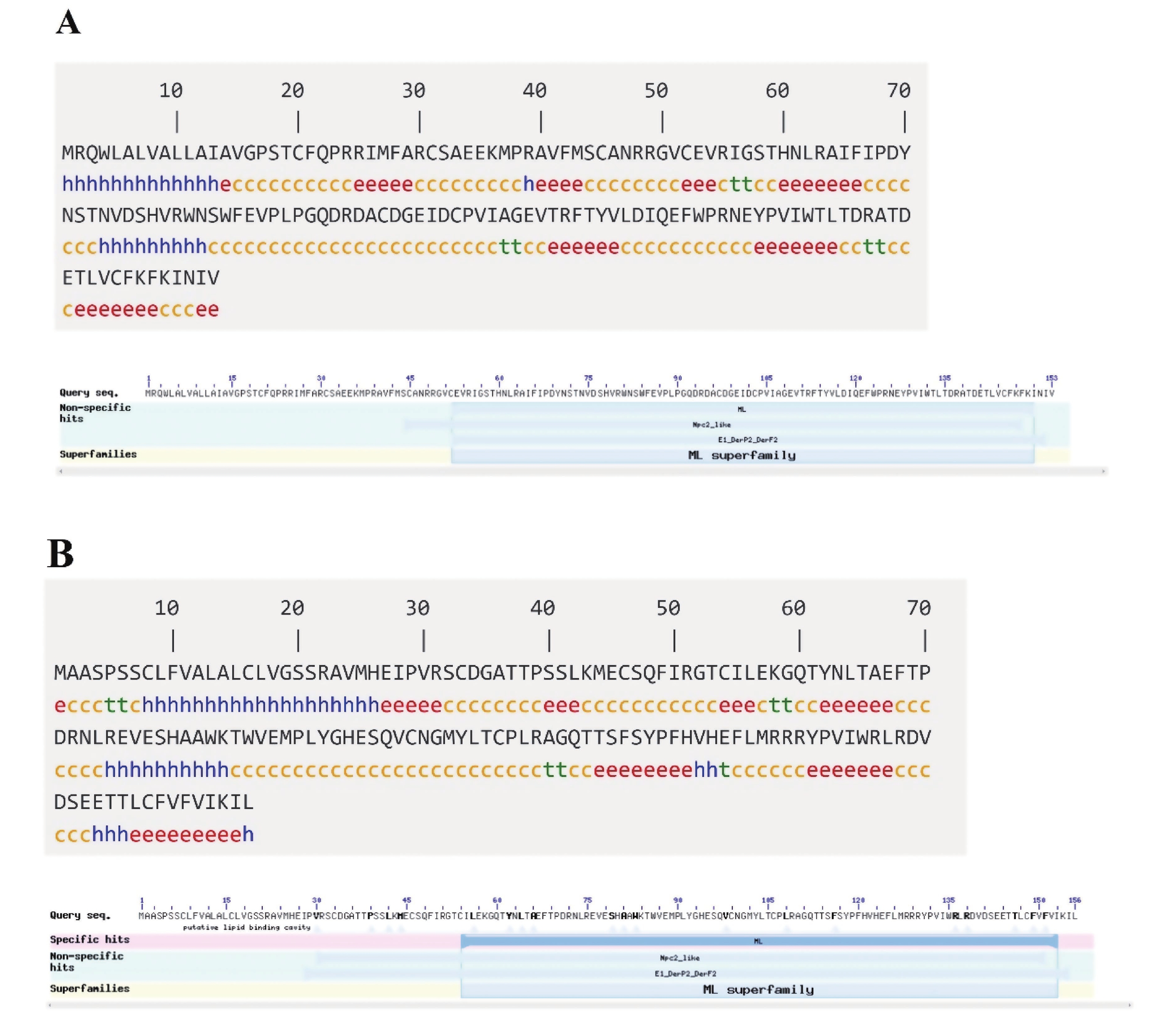
根据测序结果,拼接获得了斑节对虾2个ML基因的cDNA全序列(图1),分别命名为PmML1(GenBank:MZ291448)与PmML2 (GenBank:MZ291449)。PmML1基因由307 bp的5′非编码区(5′-Untranslated Region,5′-UTR)、462 bp的开放阅读框(Open Reading Frame, ORF)和911bp的3′-UTR组成,编码153个氨基酸;PmML2基因由95 bp的5′-UTR、471 bp的ORF和574 bp的3′-UTR组成,编码156个氨基酸。PmML1的氨基酸序列中Arg(R)、Val(V)和Ala(A)含量较高,相对分子质量为17 604.30,等电点为6.11;PmML2 的氨基酸组成中Leu(L)和Ser(S)含量较高,相对分子质量为17 719.52,等电点为6.94,说明2个蛋白可能是差异较大的小分子蛋白。PmML1的疏水平均值(GRAVY)为-0.035,为亲水性蛋白;而PmML2的疏水平均值(GRAVY)为0.024,属疏水性蛋白。2个蛋白的疏水性差异暗示其可能在不同的部位发挥生物学功能。此外,PmML1在71位发现1个“NSTN”基序的N−糖基化位点;PmML2在38和113位各有1个O−糖基化位点,在63位有1个“NLTA” 基序的N−糖基化位点。

预测显示,PmML1蛋白N端存在1个具有20个氨基酸残基的信号肽;而PmML2蛋白N端存在1个具有23个氨基酸残基的信号肽(图1)。蛋白亚细胞定位预测结果显示,PmML1蛋白定位于细胞外(包括细胞壁)、细胞质、线粒体、液泡和高尔基体的比例分别为33.3%、22.2%、22.2%、11.1%和11.1%;而PmML2蛋白定位于细胞外(包括细胞壁)、内质网和液泡的比例分别为66.7%、22.2%和11.1%。2个基因的定位差异表明他们可能在细胞内外执行不同的生物学功能。预测PmML1的半胱氨酸残基可形成3对二硫键,位点依次为31~145,45~103和52~97;PmML2半胱氨酸残基可形成3对二硫键,位点依次为33~99,46~54和106~148。这些二硫键除了用于维持PmML1和PmML2分子内构象以单体发挥作用外,可能还会形成二聚体或寡聚体、多聚体来执行某些生物学功能。蛋白二级结构分析显示,PmML1蛋白中β−片层结构占比31.37%;PmML2蛋白中β−片层结构占比为31.41% (图2)。蛋白结构域分析显示,PmML1蛋白第52~150号氨基酸残基之间有1个典型的ML super family结构域(cl00274);PmML2在第54~153号氨基酸残基之间具有1个ML结构域(smart00737)(图2)。
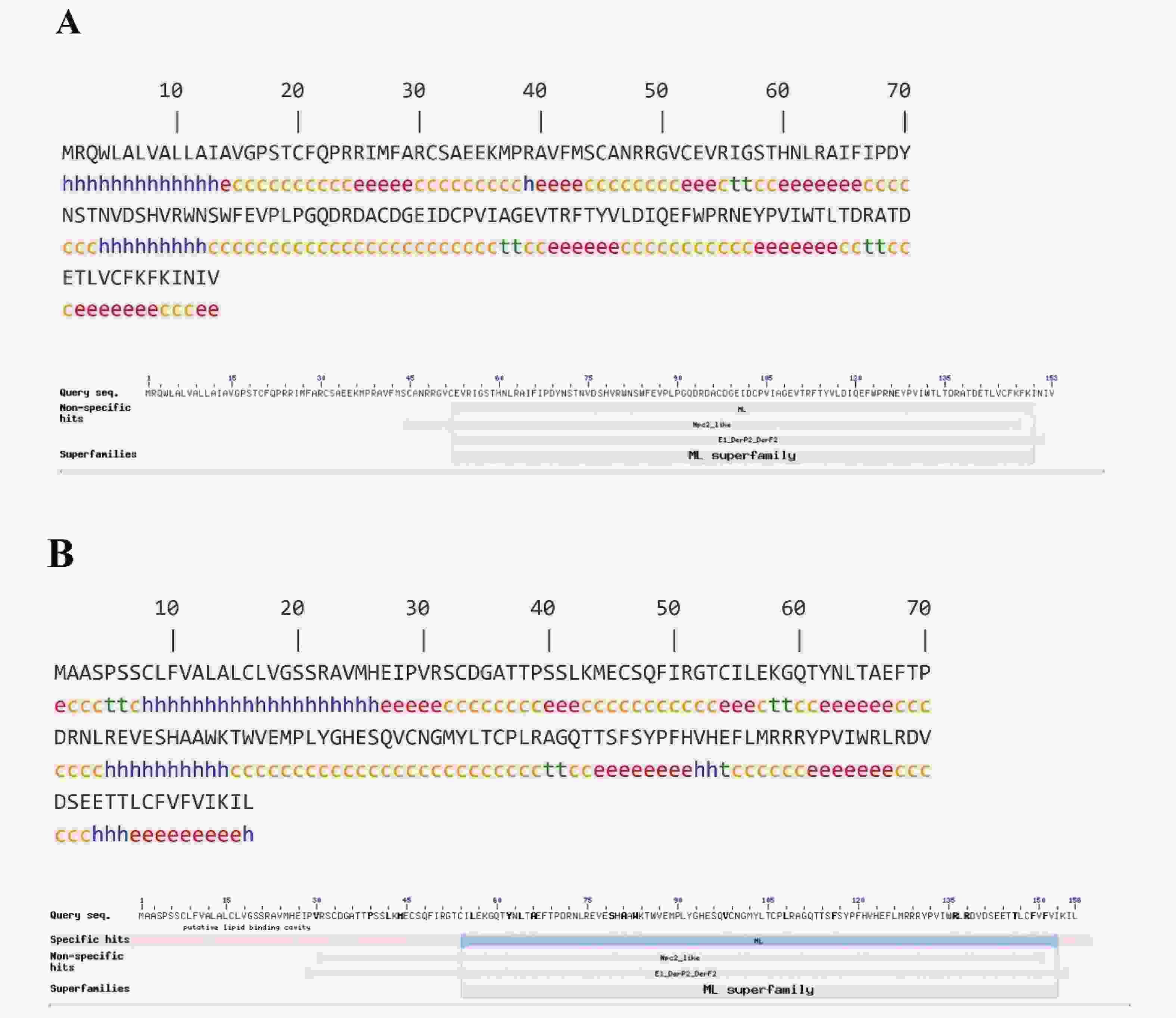

-
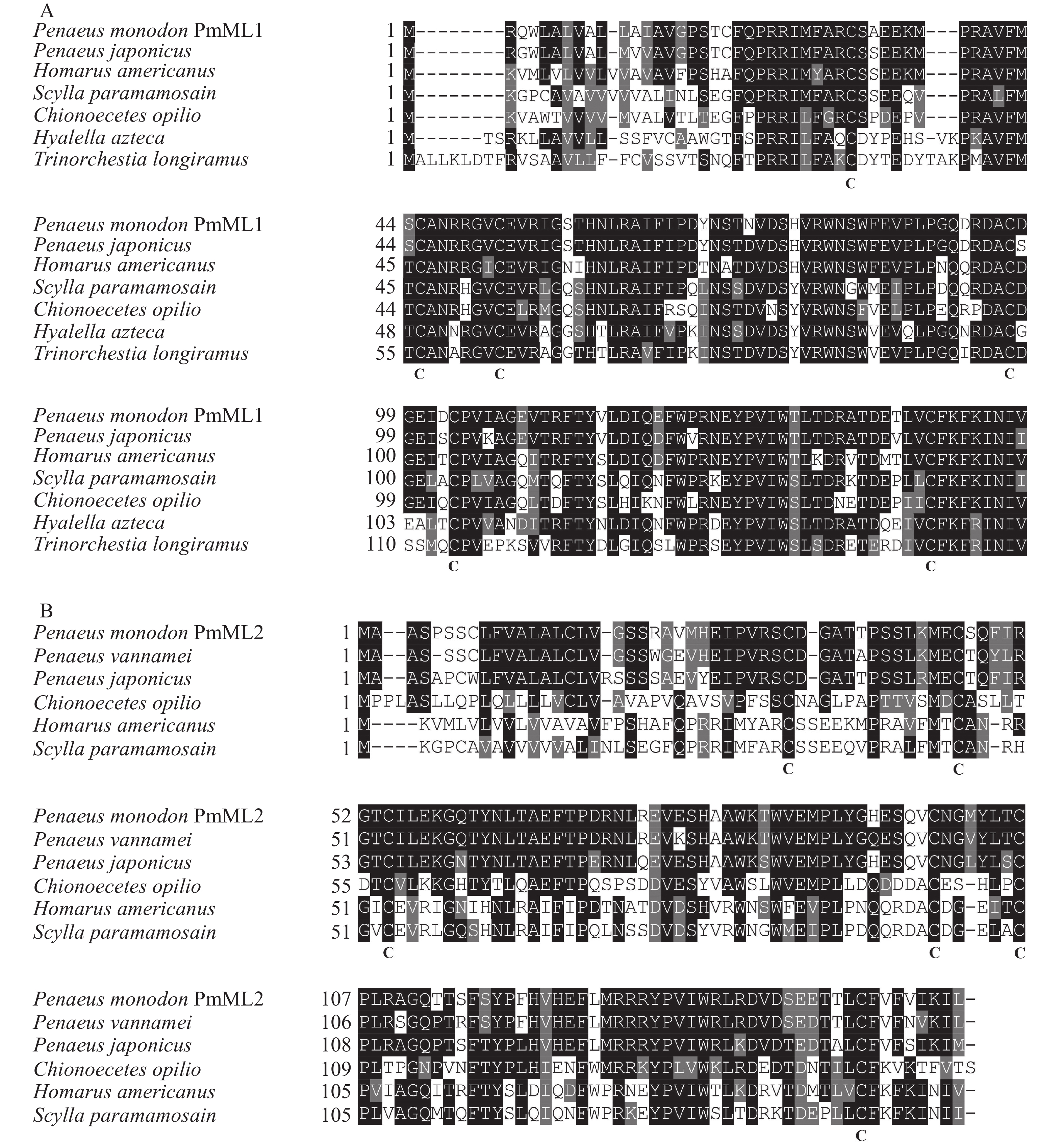
氨基酸序列比对分析发现,PmML1和PmML2之间的相似性仅为42.37%,除保守区外两者的差异氨基酸会影响中央疏水腔结构,进而识别特异性脂类,使得2个基因可能在斑节对虾生理活动中行使不同的功能。 PmML1与PmML2氨基酸序列与凡纳滨对虾、日本囊对虾等近缘动物不同ML多重比对表明其ML蛋白结构域及半胱氨酸位置相对保守(图3)。PmML1与日本囊对虾的ML蛋白MjML4相似性最高(91.5%);PmML2与凡纳滨对虾ML蛋白(XP027236046.1)相似性最高(82.8%)。系统进化分析结果显示,PmML1和PmML2分别存在于2个不同的进化分支,暗示2个基因在进化过程中相对独立,证明本研究中鉴定到的是2个不同的基因(图4)。


-
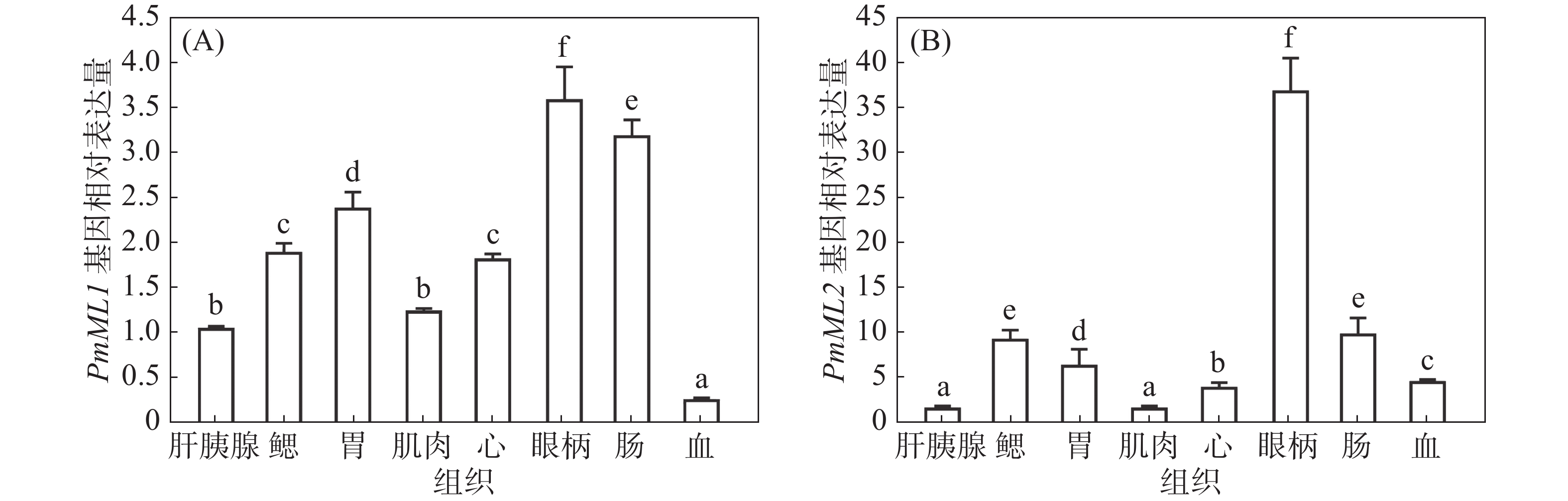
qRT-PCR检测PmML1和PmML2在斑节对虾肝胰腺、鳃丝、胃、肠、心、肌肉、眼柄和血细胞等8种组织中的表达。从图5可知,PmML1在8种组织中均有表达,以肝胰腺中表达量为对照,其在眼柄中表达量最高,其次为肠、胃、心、鳃、肌肉和肝胰腺,而在血细胞的表达水平最低。PmML2在8种组织中也均有表达,以肝胰腺中表达量为对照,其在眼柄中表达量最高,其次为肠、鳃、胃、血细胞、心、肝胰腺和肌肉,在肌肉中的表达水平最低。PmML2在眼柄的高表达,提示其可能参与了该部位重要器官X器官窦腺复合体调控的多种重要生理活动。

-
ML(MD-2-related Lipid-recognition)基因家族是由一类具有ML单结构域的蛋白质所组成,包括MD-1、MD-2、GM2激活蛋白、Niemann-Pick型C2 (NPC2)蛋白和多种广泛分布于动植物及真菌的未知功能蛋白[6]。到目前为止,已有多个物种的ML家族基因被鉴定。例如,在果蝇(Drosophila melanogaster)基因组编码8个ML家族基因,在冈比亚按蚊(Anopheles gambiae)基因组编码13个ML家族基因,在埃及伊蚊(Aedes aegypti)基因组中编码24个ML家族基因[22]。本试验利用斑节对虾2个ML相似片段,成功获得斑节对虾2个ML基因(PmML1、PmML2)的cDNA全序列,在斑节对虾中尚属首次。PmML1由153个氨基酸组成,由1个20个氨基酸残基的信号肽和3对半胱氨酸残基的ML结构域组成;PmML2由156个氨基酸组成,由1个23个氨基酸残基的信号肽和3对半胱氨酸残基的ML结构域组成,2个基因都符合ML蛋白多由140~160个氨基酸残基组成的小分子量以及都有1个N端信号肽序列和至少2对保守的半胱氨酸残基的结构特征[7]。因此,本研究获得的2个基因都是ML基因家族的新成员。
ML蛋白的ML结构域由多个β片层结构组成,通过形成2个大的β折叠构成1个Ig-like结构的中央疏水腔[23]。该中央疏水腔通过包被脂类配体的疏水部分促进其溶解性并被膜上受体和酶所识别[7]。斑节对虾PmML1和PmML2的2级结构预测显示,其β片层结构分别占比31.37%和31.41%;同源性和多重对比分析显示,在配体结合部位两者存在较高的序列变化,特别是疏水性氨基酸残基的差异。说明这两个蛋白形成的中央疏水腔的结构存在较大差异,能识别不同的特异性脂类,在斑节对虾生理活动中可能行使不同的功能。这一结果在许多研究中也得到证实,例如MD-2结合TLR4的复合体除识别LPS外,还能识别紫杉醇、人衣原体HSP60、纤维粘连蛋白的功能结构域等物质[24];而NPC2蛋白晶体结构分析则显示其内部疏水腔能结合1个胆固醇分子[25]。
ML蛋白的糖基化位点和半胱氨酸残基对ML蛋白正确行使功能发挥至关重要的作用。本研究中,PmML1在71位点有1个“NSTN”基序的N-糖基化位点;PmML2在38和113位点各有1个O−糖基化位点,在63位点有1个“NLTA” 基序的N−糖基化位点。相关研究已证实,在糖基化位点发生突变后,MD-2仍可表达于细胞表面,并保持与TLR4结合,但辅助TLR4介导NF-kb的活化能力减弱,不能促进LPS诱导IL-8分泌,也不能活化JNK途径[26-28]。在NPC2蛋白还发现不同的糖基化形式将决定其能否正确靶向溶酶体从而调节胆固醇的代谢水平[25,29]。本研究中,PmML1氨基酸序列中有7个半胱氨酸残基,除1个位于信号肽序列外,剩余6个可形成3对二硫键,PmML2氨基酸序列中有8个半胱氨酸残基,但其中有2个位于信号肽序列,剩余6个可形成3对二硫键。这些半胱氨酸残基及其形成的二硫键,将对PmML1和PmML2蛋白的分子构象及生物学功能发挥具有重要影响。半胱氨酸位点突变实验证实,MD-2分子的寡聚化不依赖任何单一的半胱氨酸残基,但95位和105位的半胱氨酸残基突变将导致MD-2丧失与TLR4结合并赋予TLR4对LPS反应的能力[28,30]。二硫键形成的共价结构,能够为许多蛋白质提供基本的稳定性并符合明确定义的几何构象[31]。MD-2蛋白半胱氨酸残基构成的二硫键可以形成分子内二硫键,也可以由游离的二硫键在分子间连接形成稳定的二聚体或寡聚体,多聚体,但单体MD-2比多聚体MD-2更易结合TLR4,能够有效促进TLR4对LPS的识别[32]。因此,笔者推测,MD-2 的二聚体、寡聚体、多聚体可能是MD-2单体避免被代谢掉的一种储存形式,但具体的分子机制有待进一步的研究。
不同的ML家族蛋白在不同物种的分布具有很大的差异。本试验利用实时荧光定量PCR技术检测发现,PmML1和PmML2基因在斑节对虾鳃丝、肝胰腺等8种组织中的相对表达量,发现PmML1和PmML2在8种组织中均有表达,PmML1在眼柄表达量最高,而PmML2在眼柄中表达量最高,且与其他组织有明显差异。总的来说,PmML1和PmML2是在机体内普遍存在的,但有组织偏好性,在眼柄中表达量最高,而在免疫组织中表达量相对较低。这在日本囊对虾和凡纳滨对虾等近缘物种的研究中也得到证实。例如,在日本囊对虾的ML家族基因中,MjML1、MjML2和MjML4在各个检测组织中均有分布,而MjML3和MjML5则仅在肝胰腺中表达[14];在凡纳滨对虾中,LvML则仅在肝胰腺组织中表达[12]。位于对虾眼柄的主要功能器官为X器官窦腺复合体(X-organ sinus gland complex, XO-SG complex),是对虾神经内分泌调节系统之一,在结构和功能上类似脊椎动物的下丘脑-神经垂体系统,它参与调控甲壳动物血糖水平、生长蜕皮和性腺发育等多种重要的生理活动[33]。斑节对虾中PmML1和PmML2在眼柄的高表达,暗示这2个基因可能主要在神经内分泌系统发挥调控作用。PmML1和PmML2在多个免疫组织表达量相对较低,暗示PmML1、PmML2 2种蛋白参与的生物学过程可能不是和免疫反应直接相关的。此外,徐伊桦等[13]对日本囊对虾MjML的研究发现,其ML蛋白可能参与了对虾的变态及蜕皮等重要生理过程。本研究的2个ML蛋白或许具有类似的生物学功能。综上所述,可以认为PmML1和PmML2是ML基因家族的新成员,可能在斑节对虾先天免疫及神经内分泌调控等方面发挥重要作用。
Identification and expression analysis of PmML1 and PmML2 in Penaeus monodon
doi: 10.15886/j.cnki.rdswxb.2022.05.003
- Received Date: 2022-01-20
- Accepted Date: 2022-05-08
- Rev Recd Date: 2022-03-31
- Available Online: 2022-05-11
- Publish Date: 2022-09-21
-
Key words:
- Penaeus monodon /
- ML (MD-2-related Lipid-recognition) /
- gene cloning /
- tissue expression
Abstract: MD-2-related Lipid-recognition proteins (ML) are a class of proteins that play an important role in the innate immunity and lipid recognition and metabolism. Two ML family genes PmML1 and PmML2 were cloned from shrimps (Penaeus monodon) by using PCR and RACE methods for bioinformatics analysis, and their expression profiles in various tissues of the shrimps were analyzed by using the quantitative real-time PCR (qRT-PCR). The results showed that the open reading frame (ORF) length of PmML1 was 462 bp, encoding 153 amino acids and that the ORF length of PmML2 was 471 bp, encoding 156 amino acids. There is a typical ML family protein structural domain in these two genes. Multiple alignments revealed that the ML domain and the cysteine position in the PmML1 and PmML2 were relatively conserved. Alignment of the amino acid sequences showed that the similarity between PmML1 and PmML2 was only 42.37%. Moreover, the similarity between PmML1 and the MjML4 of Marsupenaeus japonicus was the highest (91.5%), and the similarity between PmML2 and the ML protein of Litopenaeus vannamei was the highest (88.46%). Phylogenetic analysis showed that PmML1 and PmML2 were found in two different branches, indicating that the evolution of two genes is relatively independent, which proved that two genes were identified different. Furthermore, the qRT-PCR results showed that PmML1 expressed highest in the eye stalk, followed by in the intestine, stomach, heart, gill, muscle and hepatopancreas, and the lowest expression level was in the blood cells. PmML2 expressed highest in the eye stalk, followed by in the intestine, gill, stomach, blood cells, heart, hepatopancreas and muscle, and the lowest expression level was in the muscle. As the eyestalk is an important neuroendocrine regulatory organ, it is speculated that PmML1 and PmML2 may play a role in the neuroendocrine regulation in P. monodon. This result might provide basic data for further study of ML family genes and their biological functions in shrimps.
| Citation: | FU Danfeng, LIU Hongtao, YANG Mingqiu, HE Yugui. Identification and expression analysis of PmML1 and PmML2 in Penaeus monodon[J]. Journal of Tropical Biology, 2022, 13(5): 440-450. doi: 10.15886/j.cnki.rdswxb.2022.05.003

|
 DownLoad:
DownLoad: