
-
AP2亚族在植物的生长发育过程中起着至关重要的作用。ANT是一类重要的植物转录因子,与植物器官的发育过程密切相关,属于AP2/EREBP转录因子超家族中花卉同源基因AP2亚族,于1996年由Elliott等人从拟南芥中首次发现[1]。有研究表明,ANT基因的突变会影响拟南芥花瓣的数量,萼片的融合及花器官的位置和生长[2];ANT基因受到生长素信号的诱导影响花器官中营养物质的积累[3];ANT基因的缺失会导致植物叶片器官发育的矮小[4];ANT基因超表达会导致拟南芥各种营养器官,如叶、茎、花梗、萼片、花瓣、雄蕊、雌蕊、胚珠、果实等形态大小的增加[5];ANT基因突变会导致胚珠不正常发育以及雌配子消失[6];ANT基因促进了开花植物叶尖侧器官的生长及极性发育[7];ANT基因参与番茄花器官发育、果实初期颜色和果实大小的发育[8],参与葡萄的浆果大小[9],以及油菜、大白菜叶球的大小的发育[10]。ANT家族在众多植物生长发育中的功能已经得到证实,但在荔枝中的研究尚未见报道。荔枝(Litchi chinensis Soon.)为华南地区产量最大的热带果树之一,在经济、文化和食用价值上都具有不可忽视的地位。在荔枝实际生产过程中常因雄、雌花周期相遇短造成授粉困难,导致坐果不佳,严重影响荔枝果实产量,从而制约了荔枝产业的发展。‘妃子笑’荔枝为偏早熟品种,以肉多、味美、品相佳,产量好等优点在华南地区广泛种植,因雄花比例大,雌花比例小,易受天气影响等问题导致产量不稳定,有研究表明‘妃子笑’胚珠的发育与花型的形成密切相关[11]。随着生物技术的发展,转基因技术不断成熟,已经成为改良作物的主要手段之一[12],挖掘荔枝开花,营养器官发育相关基因,将有助于利用分子育种手段改良荔枝开花及发育过程,从而改良荔枝果实品质。本研究拟从筛选并克隆荔枝花器官相关基因ANT基因入手,通过生物信息学分析手段初步了解荔枝ANT(LcANT)的生物学特征,构建系统进化树,分析起源分化,进行组织特异性表达分析探究LcANT基因在‘妃子笑’荔枝中的表达模式,从而有助于推测ANT类转录因子在‘妃子笑’荔枝发育过程的机理,为调控‘妃子笑’荔枝品种开花结果提供分子水平上的理论基础。
-
‘妃子笑’荔枝不同组织材料均采自海南省农业科学院热带果树研究所永发基地,包括一年生根、茎、春梢(12月份采摘)、秋梢(9月份采摘)、花序、花药、果皮、果肉、种子,共9个植物组织材料。采摘后清洗所有样品,立即在液氮中冷冻,−80 ℃保存。每个材料采集3份用于重复。
-
按照植物RNA快速提取试剂盒说明书(中国华越洋生物技术有限公司)分离植物总RNA,用微量紫外分光光度计(NanoDrop 2000c,美国赛默飞)测量RNA浓度与质量,OD260 / OD280吸收比(1.9~2.0),并通过在1.0%琼脂糖凝胶中的电泳鉴定RNA的完整性。使用约1 μg总RNA,1 μL Oligo(dT)引物和1 μL反向逆转录酶(200 U·μL−1,Takara,日本)在20 μL反应混合物中参照反转录试剂盒(宝生生物工程有限公司,大连)说明,合成第一链cDNA。
-
参照本课题组荔枝基因组数据库,通过Primer 5.0 进行cDNA全长引物设计,上游引物为5′- GCACCGAGAGATACTGCTAAGT -3′,下游引物为5′- CTGGCAAGTGTGACATGGAGAT-3′,委托上海生工生物合成有限公司进行引物合成。以提取的‘妃子笑’cDNA为模板,根据普通PCR试剂盒说明,进行50 μL PCR反应体系扩增全长,扩增体系如下:5×buffer 5 μL,10 mmol·L−1 dNTPs 1 mL,上下游引物各1 μL,模板5~100 ng,F8高保真酶1μL,无菌无酶水补至 50 μL;反应程序为:95 ℃预变性10 min,95 ℃持续30 s,60 ℃退火2 min,72 ℃延伸30 s,40个循环,72 ℃维持10 min,4 ℃保存。
通过在1.0%琼脂糖凝胶中的电泳鉴定DNA的完整性和准确性,并将单一条带电泳产物切胶回收、纯化后与T载体(PMD18-T)连接。T载体的连接体系为:PMD18-T Vector 1μL,目的片段 1μL,dH2O 3 μL,Solution 5 μL;16 ℃反应30 min。T载体的连接体系全量10 μL加入到已提前冰上融化的100 μL大肠杆菌感受态(DH5α)中,继续冰上放置30 min,42 ℃热激45 s,再置于冰上1 min,加入890 μL LB液体培养基,37 ℃,180 r·min−1振荡培养60 min,将 100 μL菌悬液涂布于含100 μg·L−1氨苄青霉素的LB固体培养基上,37 ℃倒置培养12 h。挑取3~4个白色菌落加 1 mL LB液体培养基于 25 ℃、180 r·min−1条件下振荡培养8~12 h,将菌液进行PCR扩增,PCR体系为 20 μL的混合液(1 μL菌液,上下游引物各1 μL,10 μL 2×Master Mix,dH2O补足至20 μL)。
-
通过GSDS2.0 (
http://gsds.cbi.pku.edu.cn/index.php )进行内含子外显子分析;通过NCBI的CDD (http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi )数据库进行蛋白质保守结构域预测;通过protparam (https://web.expasy.org/protparam/ )进行蛋白质分子量和等电点预测;通过ProtScale (https://web.expasy.org/protscale/ )进行蛋白质亲水疏水分析;通过PSORT II (https://www.genscript.com/psort.html )进行核定位信号的预测;通过softberry (http://linux1.softberry.com/all.htm )进行蛋白质亚细胞定位;通过SOPMA (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html )进行蛋白质二级结构的预测;通过NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi )与其他植物进行BLASTP同源性比对;通过MEGA7.0软件构建系统发育树;通过DNAMAN8软件进行克隆基因的序列比对。 -
通过Primer 5.0 进行设计,上游引物为5′- GCTTCTGCCTCCGTTGGTTCCAT -3′,下游引物为5′-TCTGCCTTGGAGCCGTGACACA-3′,使用SYBR Green实时荧光定量试剂盒在QuantStudio 6 Flex荧光定量PCR仪(American)上进行实时定量PCR(q-PCR)。该反应系统的体系和扩增参数如下:SYBR Green10 μL,上下游引物各0.4 μL,模板2 μL,无菌无酶水7.2 μL。在95 ℃预变性10 min,然后在95 ℃持续30 s,55 ℃退火30 s和72 ℃持续30 s,40个循环。肌动蛋白基因Actin作为内参基因[13],上游引物5′- TGCATGAGCGATCAAGTTTCAAG -3′,下游引物5′- TGTCCCATGTCTGGTTGATGACT -3′[14],所有反应重复3次。Ct值通过QuantStudioTM实时PCR软件以默认设置进行测量。基因的相对表达水平通过2−ΔΔCt方法计算[15]。Q-PCR用于验证Litc12A14880在 9种‘妃子笑’荔枝9种营养器官组织中的相对表达水平。
-
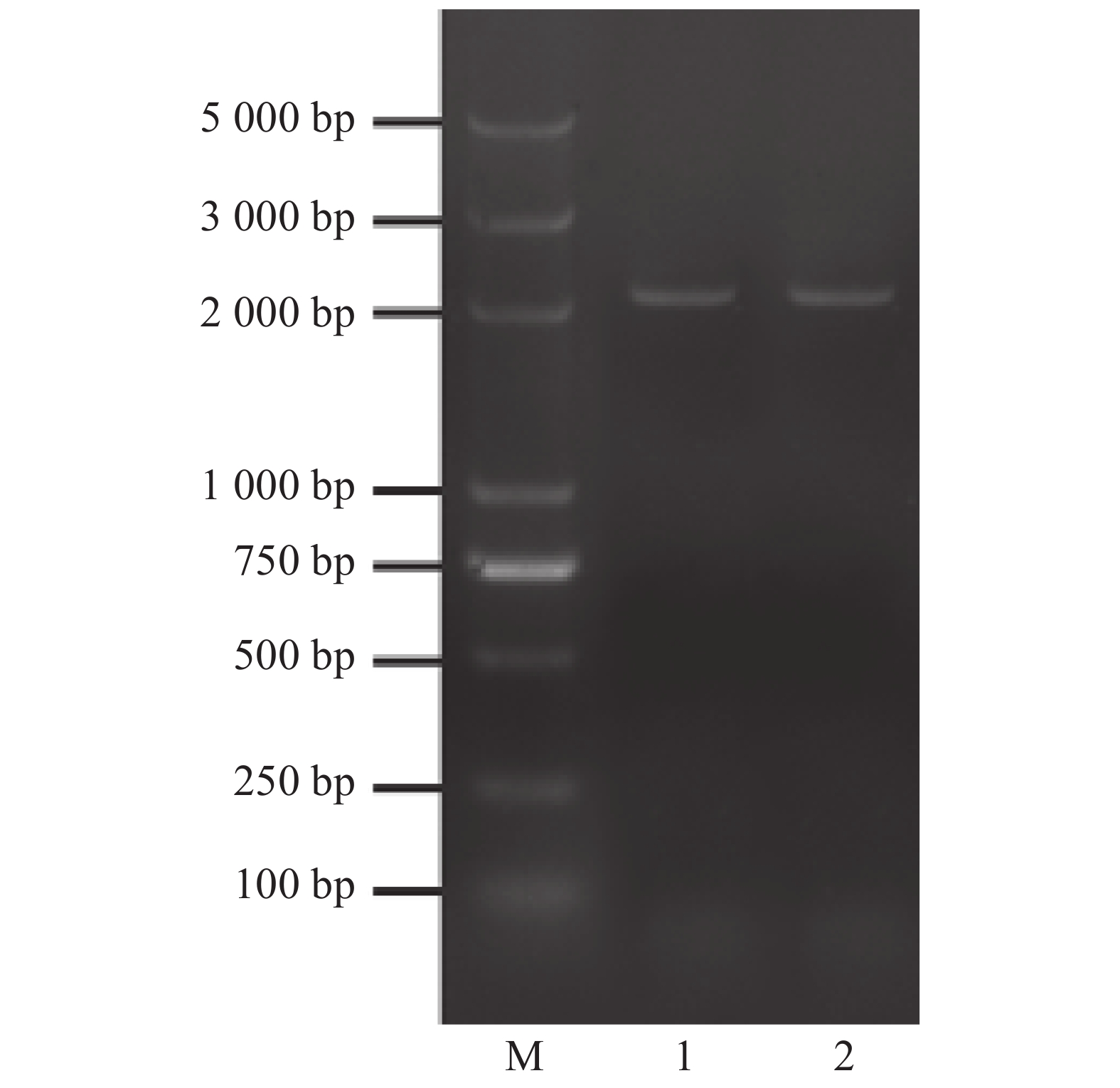
根据课题组的前期转录组数据设计了扩增cDNA全长的引物12AF和12AR,进行荔枝LcANT基因的克隆,PCR产物在0.9%的琼脂糖凝胶上进行30 min电泳,在2 000 bp处观察到目的条带(图1),将目的条带进行切胶回收,进行TA克隆,将阳性克隆菌液送测序公司进行测序。该基因基因组全长5 331 bp,cDNA编码区全长为2 087 bp,与转录组数据进行序列比对(图2),发现测序结果在序列的5′段43 bp左右,和3′端45 bp左右与转录组数据有所不同,可能是因为基因序列较长,测序结果误差导致,此段序列以转录组数据结果为准,此外,测序结果中间段也与转录组数据有少许不同,经过3次阳性菌落重复测序,结果均一致,推测序列中间段误差可能是由于转录组数据并不完全准确。目的序列与转录组匹配度高达99.71%,可认为成功克隆了荔枝LcANT基因。
Figure 1. Cloning of LcANT from Litchi

Figure 2. The alignment of LcANT sequences
-
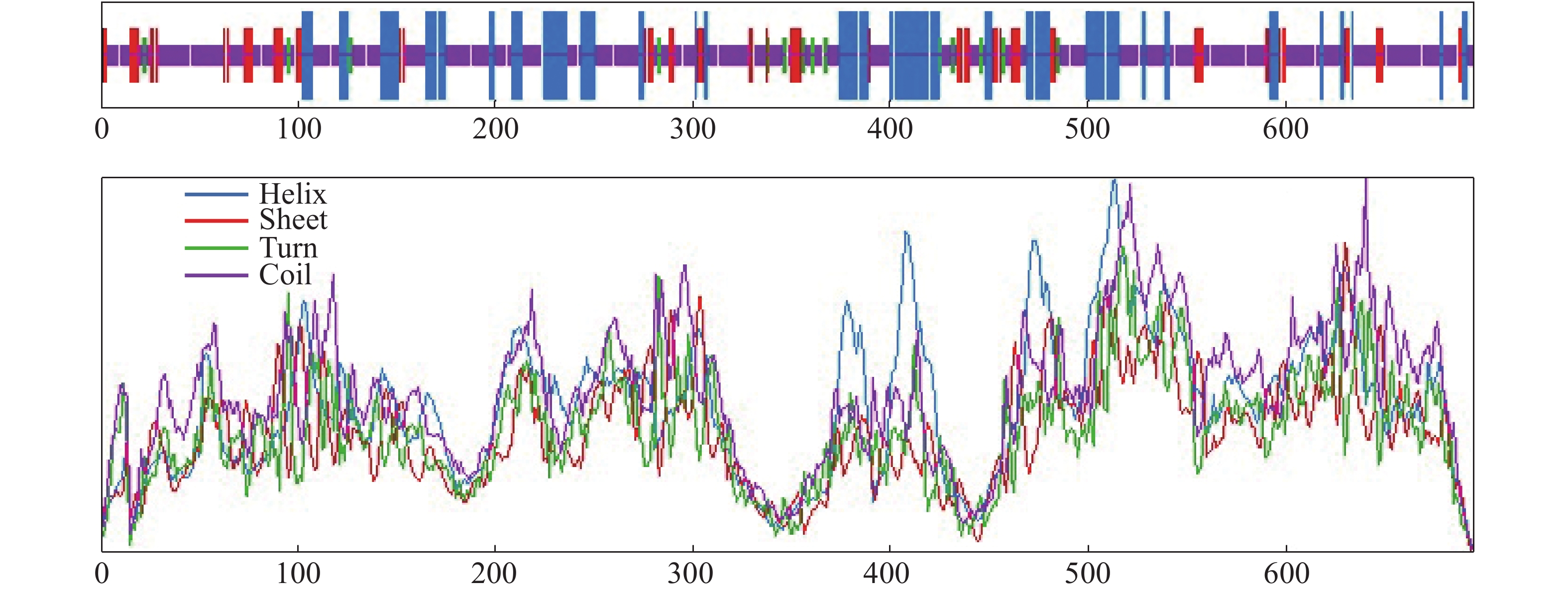
LcANT基因具有7个内含子(图3),与拟南芥ANT类转录因子基因结构内含子个数相似,表明AP2亚组在长远的历史进化中高度保守。该基因共编码695个氨基酸,在337~406位置和437~500位置有2个保守的AP2/EREBP结构域(图4),每个AP2/EREBP保守结构域上都具有11个DNA结合位点,这些位点作为AP2/EREBP结构域的特征性结合位点,与GCC-box顺式作用元件结合,启动植物乙烯应答反应、参与植物应激反应调节及植物生长发育的调控等[16],此外,在362~395位置还有1个包含31个氨基酸残基的核定位信号(NLS),该序列为非典型核信号序列,通过与核蛋白相互结合完成LcANT蛋白向细胞核内的转移[17]。

Figure 3. The structure of LcANT gene

Figure 4. Analysis of conserved domain of LcANT protein
-
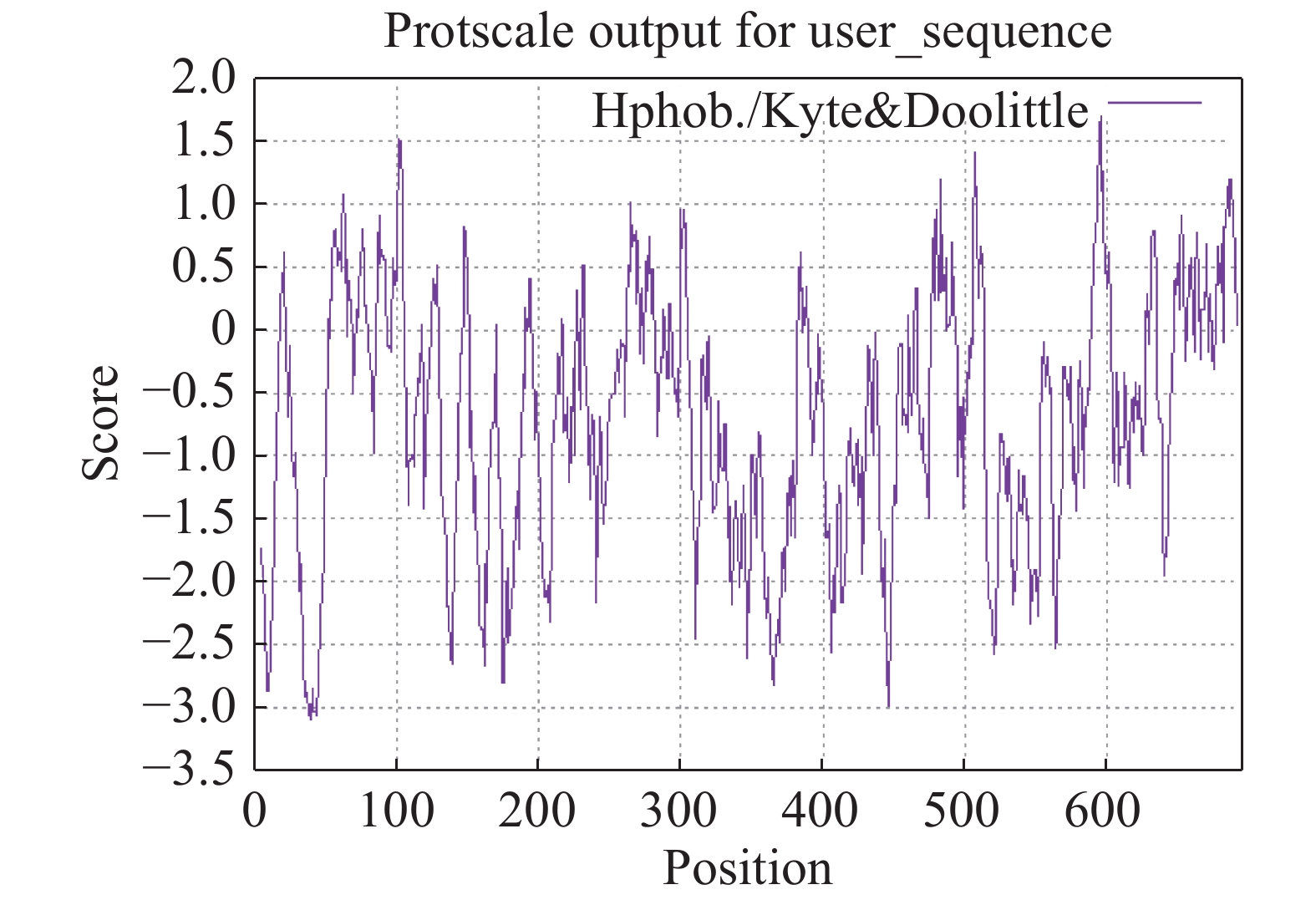
LcANT蛋白相对分子量为76.57 kDa,等电点为6.34,亚细胞定位预测其定位于细胞核中。从亲水性分析预测结果来看,LcANT蛋白质多肽链中包含亲水性氨基酸和疏水性氨基酸,其中最大值为1.71,最小值为−3.09,整体来看,疏水性氨基酸明显多于亲水氨基酸,且分值更大,更具有疏水性,因此LcANT蛋白质属于疏水性蛋白(图5),这更有助于结合其他蛋白发挥调控及运输功能作用。通过对LcANT蛋白进行二级结构预测,LcANT蛋白质二级结构不同类型占比分别是:α−螺旋23.02%,β−折叠12.52%,β−转角3.74%,无规则卷曲60.72%(图6)。发现LcANT蛋白质二级结构不同于其他花发育基因蛋白,例如参与花发育的MADS-box蛋白的二级结构主要由60%的α−螺旋构成,而LcANT蛋白的二级结构中的比例高达60%的结构类型是无规则卷曲,而α−螺旋的比例则明显低于大多数花发育相关基因[18]。
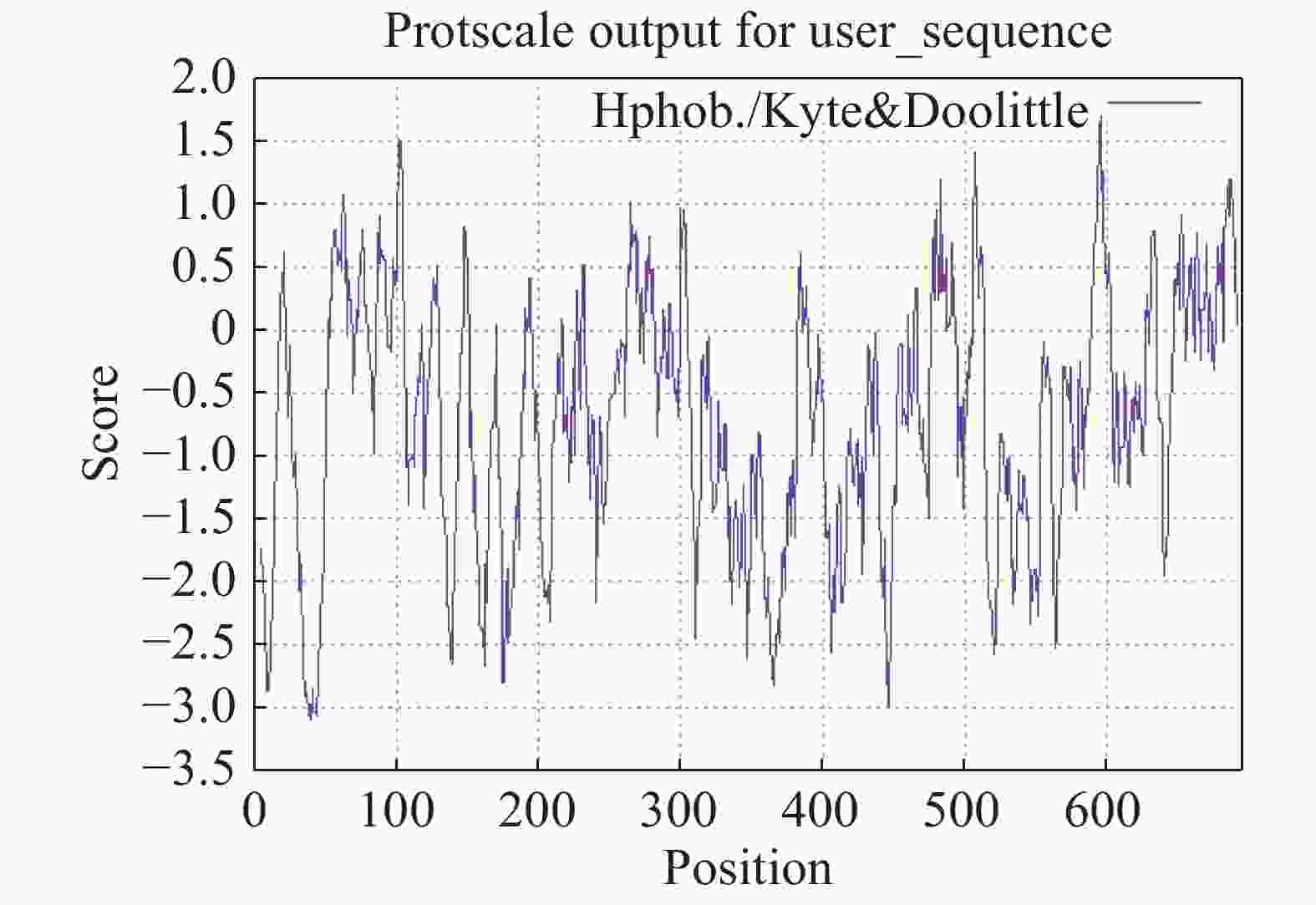
Figure 5. Hydropathy analysis of LcANT protein
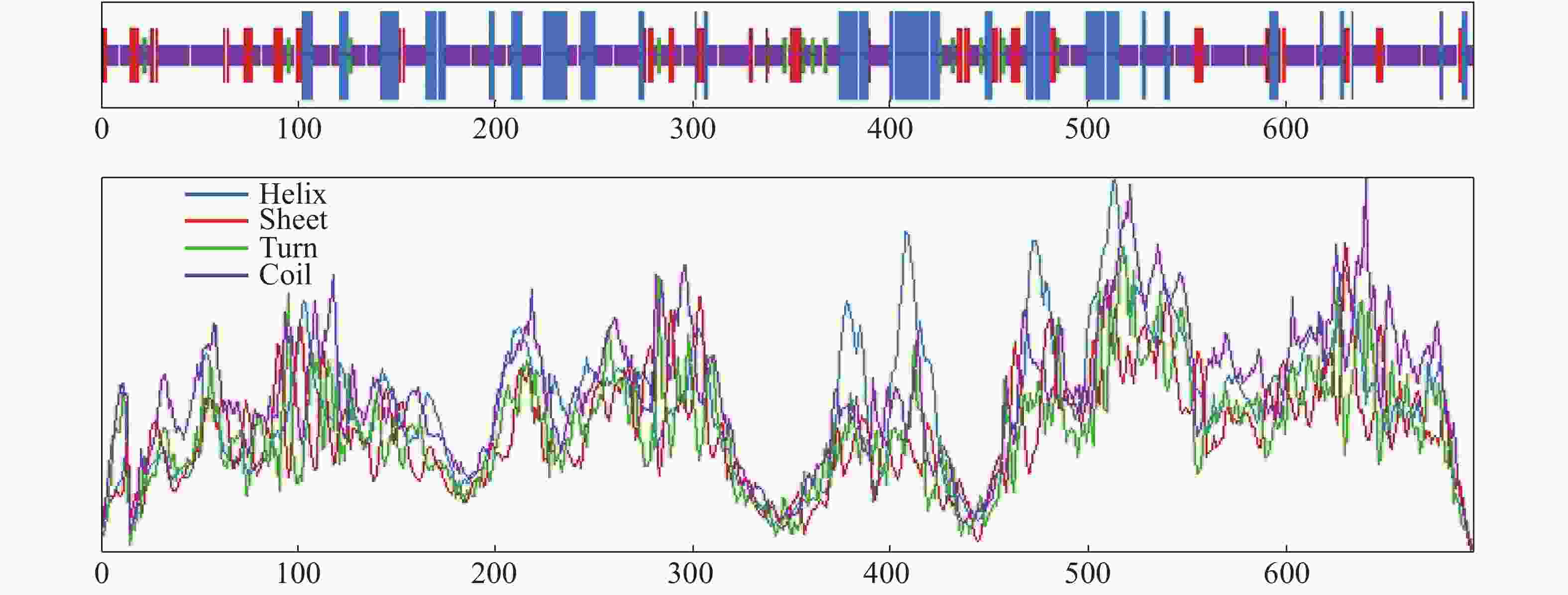
Figure 6. Prediction of secondary structure of LcANT protein.
-
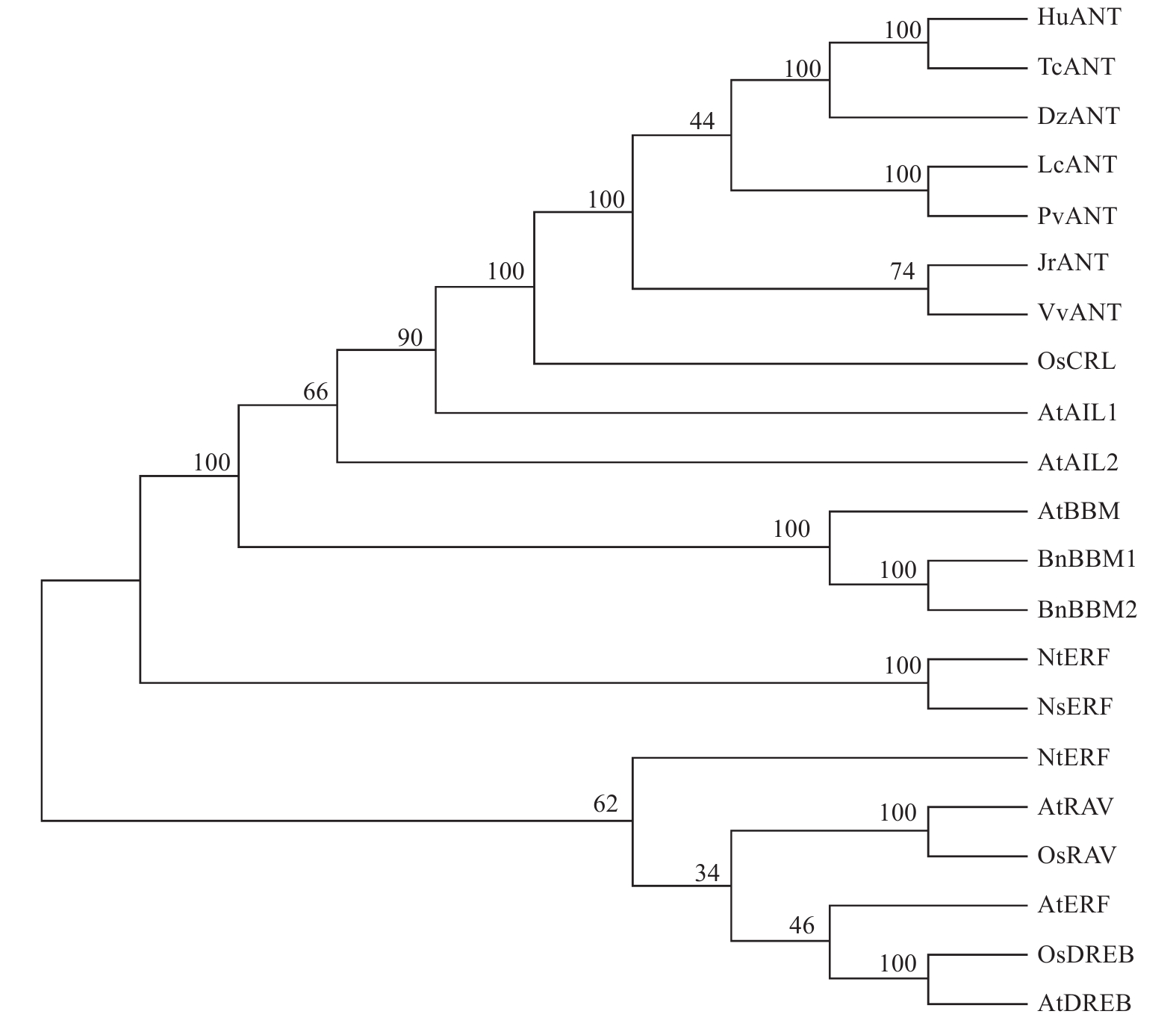
LcANT属于AP2/EREBP转录因子家族的AP2亚族成员。根据2018年丁谦等人的AP2转录因子结构分类[19],AP2亚族可细分为euAP2亚组和ANT亚组,可将LcANT划分为ANT亚组。为了进一步确定LcANT的身份,将LcANT蛋白序列与其他物种的AP2、ERF、DREB、RAV蛋白进行序列比对,并通过MEGA7.0软件以NJ算法进行1 000次自举重复构建系统发育进化树(图7)。结果显示,LcANT蛋白与AP2家族蛋白聚集到1个分支,于ANT亚组距离较近,而与AP2其他亚组成员,如AIL、BBM等距离较远,与ERF亚族、DREB亚族、RAV亚族等距离更远。除此之外,荔枝LcANT蛋白与木本植物开心果PvANT蛋白亲缘最近,其次是榴莲DzANT、可可TcANT及草本植物锦葵HuANT。
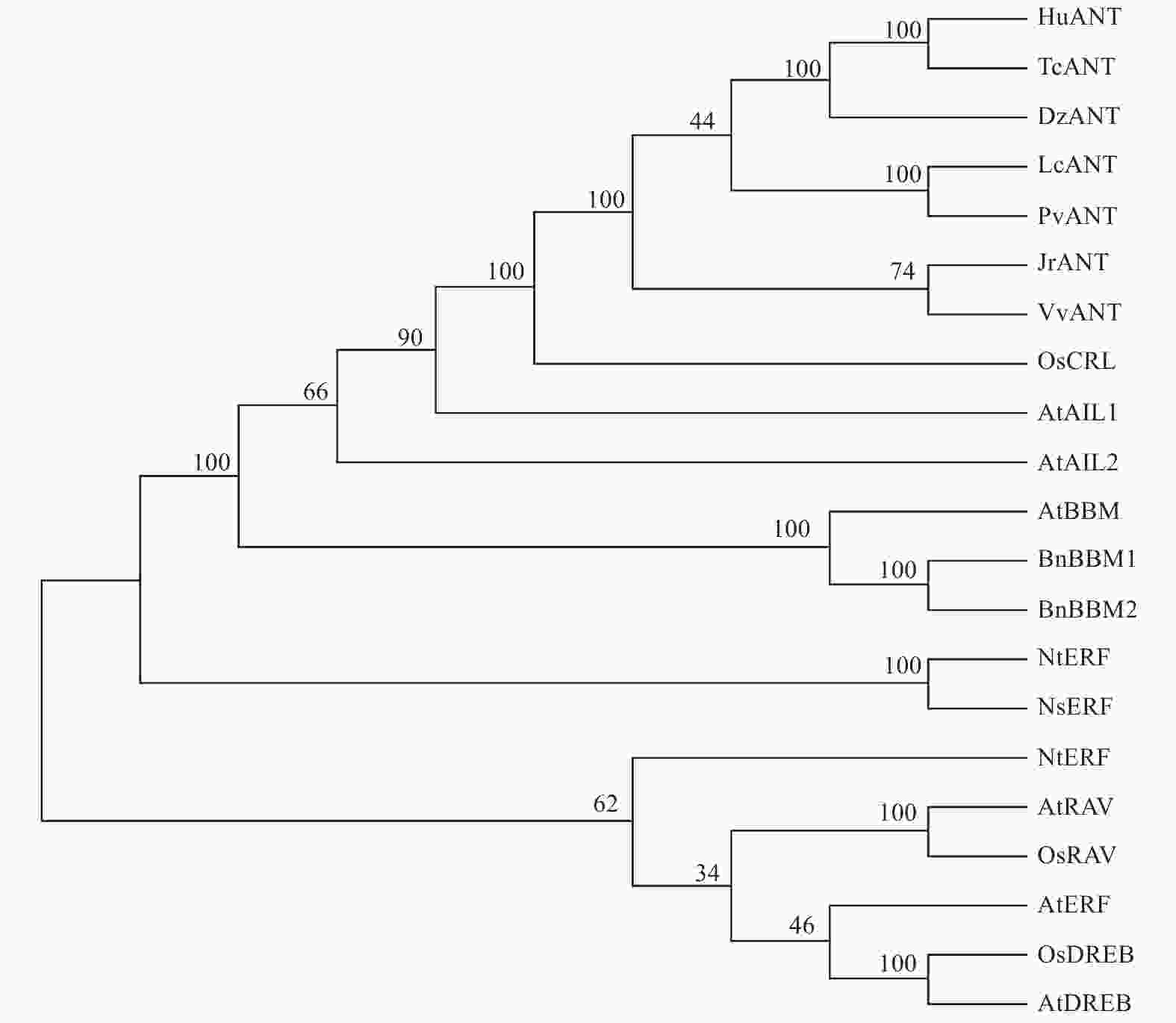
Figure 7. Phylogenetic relationship of LcANT with ANT of other plant species
为了弄清LcANT蛋白与众多植物ANT转录因子氨基酸序列的相似度,从NCBI中选取了8个物种的共10个ANT蛋白(ANT-Pistaciavera (XP_031273069.1),ANT-Herraniaumbratica (XP_021284854.1),ANT-Theobromacacao (XP_007018428.2),ANT-Duriozibethinus (XP_022766436.1),ANT-Durio zibethinus (XP_022772252.1),ANT-Juglans regia (XP_018810432.1),ANT-Vitis vinifera (RVX22674.1),ANT-Vitis vinifera (XP_002285467.2),ANT-Ricinus communis (XP_002510048.),ANT-Quercus suber (XP_023872648.1))进行氨基酸序列比对(图8),结果显示,LcANT蛋白与众多植物ANT转录因子的氨基酸序列具有高度相似性,其中与开心果的同源性最高,高达82%,与哥伦比亚锦葵同源性为79%,与可可、葡萄、榴莲、胡桃、欧洲栓皮栎、荨麻的同源性分别为79%,78%,77%,76%,76%,75%,并且在2个AP2/EREBP保守结构域范围337~406,436~500氨基酸残基位置也完全保守。由此说明,LcANT蛋白为荔枝AP2亚族中的ANT亚组成员。

Figure 8. Multiple alignments between LcANT sequences and other ANT sequences
-
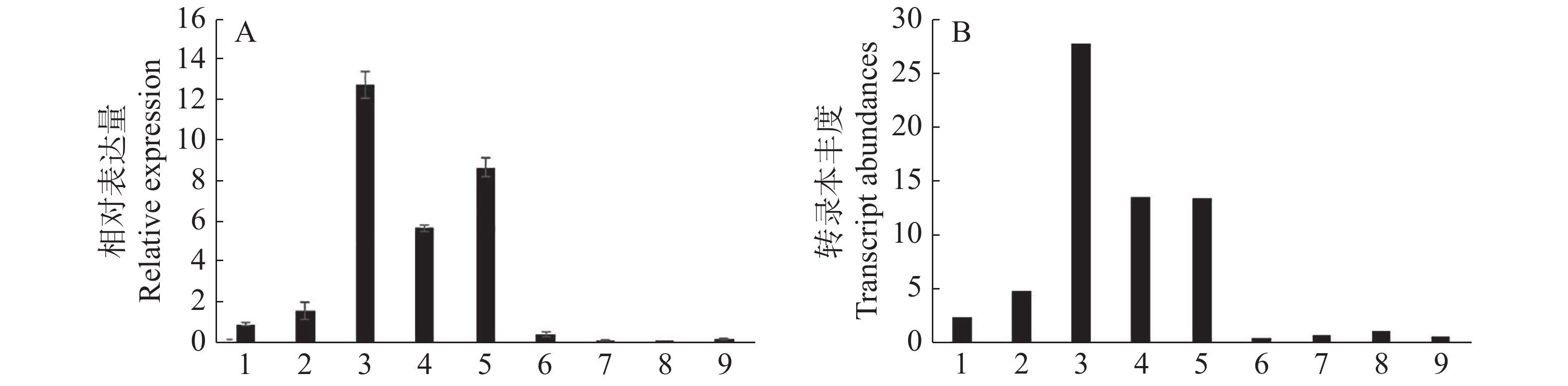
植物体不同器官和组织的生长发育过程受到多种基因的调控和协同作用,荔枝花器官发育也不例外。以肌动球蛋白Actin为内参基因,通过q-PCR分析LcANT基因在‘妃子笑’荔枝9个不同组织(根、茎、春梢、秋梢、花序、花药、果皮、果肉、种子)的相对表达量,并绘制柱状图(图9A)。结果显示,LcANT基因在大多数组织中都有不同程度的表达量,其中在春梢、秋梢和花序中相对表达量较高;而相对于叶片组织,在春梢中相对表达量较秋梢更高;相对于其他组织,在花器官中的表达量较其他组织更高。以上结果与转录组数据趋势基本一致(图9B)。
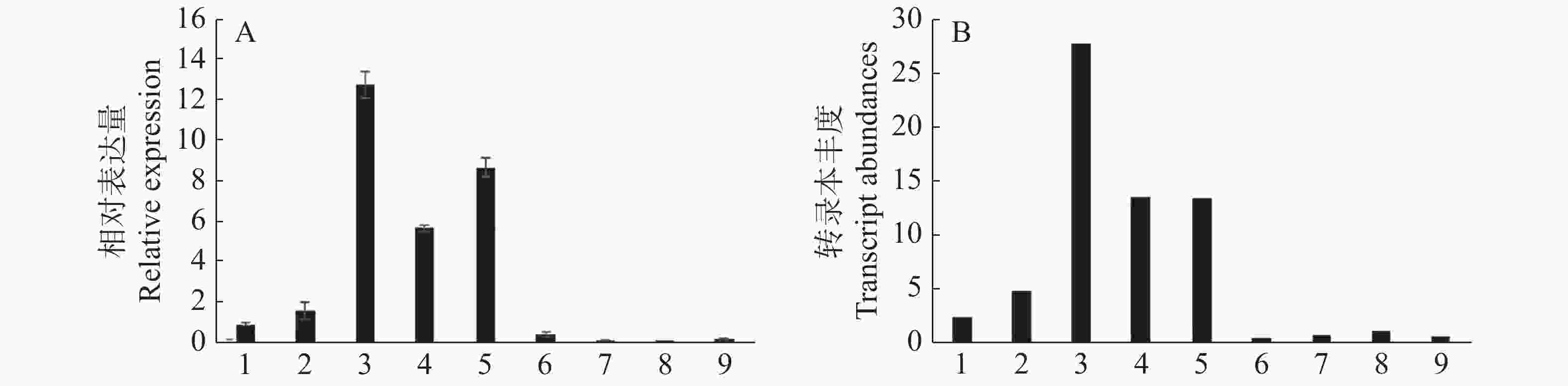
Figure 9. Relative expression and transcript abundances of LcANT in different organs of litchi
-
AP2/EREBP转录因子是植物中最大的转录因子家族之一,迄今为止,研究过的AP2亚家族的成员均在植物细胞增殖和分化方面起到重要作用[20]。而ANT作为AP2亚组的分支之一,参与植物生长发育的精确调控[21],ANT基因不仅在分生旺盛组织如腋芽中表达,还在胚珠、子房、花等营养器官中表达,甚至还在根中表达,说明整个植物生命周期的发育都离不开ANT基因的参与。ANT基因的参与方式为可通过与其他转录因子如BBM,AIL等相互协同促进响应生长激素的LFY上调表达,从而提前植物开花时间[22];有可能抑制生长素信号传导的阻遏物ARF2基因的表达,从而延缓植物叶片衰老[23];也有可能作用于靶标之一的cyclinСYCD3∶1基因[24],调节植物细胞的分裂过程;还有可能通过异位转录使ARGOS(一种生长素诱导型编码跨膜蛋白的基因)表达增强,从而控制植物器官的大小[25]。
荔枝LcANT基因cDNA长2 088 bp,编码695个氨基酸,包含2个由60~70个氨基酸残基构成的AP2/EREBP保守结构域,这些结构域上的结构位点可特异性识别并结合GCC-box顺式元件,精准调控植物对内外环境的响应。荔枝LcANT蛋白序列上包含1个非典型核定位信号并且预测该蛋白定位于细胞核中,表明LcANT基因可能在细胞核中实现对下游基因转录水平的调控[26],且其编码蛋白属于疏水性蛋白,蛋白侧链可能具有更多的疏水性残基,而这些残基往往为蛋白质的活性位点,可结合其他分子或基团,促进LcANT作用的发挥。另外,LcANT蛋白质二级结构中无规则卷曲构象类型比例较多,卷曲的柔性构象可使肽链改变走向,更利于与结构相对刚性的α螺旋和β折叠进行连接,形成多样化的蛋白质三级空间结构,揭示了该蛋白在植物体中可能发挥着多种功能作用[27]。这与荔枝植物基因组中的大部分ANT基因结构和性质是类似的。通过系统发育分析可见,荔枝LcANT蛋白与许多物种中的AP2亚家族成员亲缘距离较近,与AP2亚族的ANT亚组基因都具有高度同源性,表明由于蛋白质结构相似,ANT转录因子可能在这些物种之间发挥相同或相似的作用,说明这些物种可能源于同一祖先。此外,这些不同物种间也存在差异性,在植物繁衍过程中经过数百万年的进化,为适应不同环境生存进化出特有的基因组,这些基因通过外界信号刺激调控网络,为运应不同生物及非生物的胁迫,从而产生了功能的分化[28]。春季长出的新梢称为春梢,分裂期较旺盛,营养物质较丰富,夏季后长出的新梢称为秋梢,由于夏季炎热,生长受到抑制。q-PCR结果表明,在叶片组织中,LcANT基因在荔枝春梢中丰度更高,说明LcANT基因在植物组织细胞分裂生长的阶段发挥一定的作用,可能是通过调控生长因子和细胞周期因子来促进细胞分化,增加细胞数量和大小,从而调控营养器官的生长和发育;此外,LcANT基因在花序中丰度较其他组织(根,茎,花药,果肉,果皮,种子)高,表明ANT家族在植物花发育中起关键作用。兰花[29],菊花[26],红掌[30],拟南芥[31]中的研究也有类似的现象,例如在ANT基因可与MADS-box基因协同控制兰花的颜色,拟南芥ANT和AIL转录因子可与B类转录因子并行作用于拟南芥花卉器官的萌发,形状和性状的改变。花器官发育的ABC模型决定了花发育的过程。荔枝LcANT究竟在荔枝花发育的哪个阶段起作用,以及与A,B,C3类基因如何发挥作用,还有待进一步研究。
本研究成功克隆了荔枝LcANT基因,分析了荔枝LcANT基因的结构及蛋白性质,并确定了荔枝Litc12A14880基因参与调控荔枝花器官及幼嫩组织的生长发育,后期将进一步确定基因功能,为后续研究LcANT基因的具体功能及其他ANT亚组成员提供分子水平理论基础。
Cloning and Expression Profiling of an AP2 Homologous Gene of Litchi ‘Feizixiao’
DOI: 10.15886/j.cnki.rdswxb.2020.02.009
- Received Date: 2020-02-13
- Rev Recd Date: 2020-03-03
- Available Online: 2020-07-03
- Publish Date: 2020-06-01
-
Key words:
- Litchi chinensis Sonn. /
- ANT transcription factor /
- expression analysis
Abstract: An AP2 homologous gene LcANT was cloned from Litchi ‘Feizixiao’ (Litchi chinensis Sonn. Feizixiao) based on the litchi whole genome database available from the Environment and Plant Protection Institute. The gene LcANT was 2 087 bp in full-length cDNA, encoded 695 amino acids and contained two AP2/EREBP conserved domains. Bioinformatics analysis showed that the LcANT contained 7 introns, and that the subcellular localization was predicted in the nuclear. The LcANT protein was hydrophobic and its secondary structure wass mostly random coils. BLAST sequence alignment analysis showed that the LcANT had a high homology in amino acid sequences with the ANT transcription factors from other plants, and had a maximum identity of up to 82% with the ANT from pistachio (Pistacia vera). The q-PCR was used to characterize the expression pattern of the LcANT in the tissues of Litchi 'Feizixiao', and the results showed that the LcANT had higher expression levels in spring shoots, ovary, inflorescence, and flower buds, implying that this gene may regulate the growth and development of floral organs and other plant vegetative organs. The acquisition of the LcANT gene provides a molecular basis for further research focused on its function on the growth and development of vegetative organs of Litchi.
| Citation: | Men Jiali, LI Fang, SUN Jinhua, XU Yun. Cloning and Expression Profiling of an AP2 Homologous Gene of Litchi ‘Feizixiao’[J]. Journal of Tropical Biology, 2020, 11(2): 190-199. doi: 10.15886/j.cnki.rdswxb.2020.02.009

|
 DownLoad:
DownLoad: