-
木薯(Manihot esculenta)被称为“热带奇迹”,为多年生灌木,被誉为“淀粉之王”。作为缓解全球贫困、饥饿与营养不良问题的核心作物之一,木薯具备极高的基础光合效率,且能在极端恶劣的生长环境下维持存活,这一特性使其成为保障粮食与营养安全的重要潜力作物[1−3]。而木薯细菌性枯萎病(cassava bacterial blight,CBB)是由黄单胞菌(Xanthomonas axonopodis pv. manihotis,Xam)所引起的危害木薯的重要病害之一[4],对木薯产业发展具有极大危害。另外,栽培种木薯的氢氰酸积累量显著低于野生木薯[5]。已有研究显示,在木薯中抑制生氰糖苷生物合成的关键限速基因CYP79D1和CYP79D2的表达,会改变叶片和根部的氰苷合成水平[6]。另外CYP71E7是催化氰苷(亚麻苦苷和百脉根苷)生物合成的关键步骤,是该合成途径不可或缺的关键基因,但CYP71E7是否参与调控木薯对细菌性枯萎病的抗性,目前尚未不清楚。
细胞色素P450(cytochrome P450,CYP450)属于单加氧酶类[7],广泛参与植物各类初生及次生代谢进程,对植物的生长发育与胁迫中扮演着重要角色[8]。植物CYP450可归类为11个家族簇,分别涵盖7个单家族簇(CYP51、CYP74、CYP97、CYP710、CYP711、CYP727、CYP746)和4个多家族簇(CYP71、CYP72、CYP85、CYP86),其中家族最大、基因数量最多的是CYP71亚家族,占总量的50%以上[9]。CYP71亚家族基因的表达受光照、温度、机械损伤及病原菌侵染等多种环境因子调控[10]。在水稻(Oryza sativa)中过表达CYP71Z2可持久且稳定的增加植株对细菌性枯萎病的抵抗力[11]。CYP71家族基因通过直接或间接调控倍半萜类植保素的合成和代谢,从而抵御病虫害的入侵。拟南芥(Arabidopsis thaliana)中异源表达海岛棉GbCYP86A1-1提高拟南芥对黄萎病的耐受性[12]。高粱(Sorghum bicolor)的CYP79A1和CYP71E1—可在烟草(Nicotiana tabacum)和拟南芥中表达,形成新型氰化植物[13],而高粱中所产生的含氰苷对害虫有毒性[14]。此外,杨树(Populus)中的CYP79D6v3和CYP79D7v2基因参与醛肟等挥发性次生代谢物的合成,这类化合物不仅可以驱赶植食性昆虫,可能直接参与植物的防御调控网络[15]。
本研究从木薯中筛选并成功克降得到CYP71基因家族的成员MeCYP71E7,对其开展生物信息学分析与功能初步验证,旨在揭示MeCYP71E7在木薯抗病木薯抗细菌性枯萎病过程中的潜在功能,为后续深入解析木薯抗病机制提供关键基因资源,同时也为木薯的抗性遗传改良提供理论参考。
-
供试木薯品种为‘华南124’,选取健康木薯茎秆剪切为长度10~15 cm的节段,每段保留2~3个完整芽点,随后定植于装有混合基质(V营养土︰V蛭石= 2︰1)的花盆中,将培养条件设置为温度28~30℃,光周期为12 h光照/12 h黑暗。本实验所用烟草为实验室长期保藏的本氏烟草(Nicotiana benthamiana)。
在Phytozome14数据库(https://phytozome-next.jgi.doe.gov/)中检索获取MeCYP71E7的编码序列(coding sequence,CDS),借助Primer5.0完成引物设计,引物的合成与后续测序服务均委托深圳华大基因科技有限公司提供,所有引物相关参数详见表1。
引物名称
Primer引物序列(5′−3′)
Primer sequence(5′−3′)pCAMBIA2300-MeCYP71E7-F CGCGGATCCATGTCCGTAGCCATTCTAAC pCAMBIA2300-MeCYP71E7-R ACGCGTCGACATCCCATTTGTGTTTTTTAG pGADT7-MeCYP71E7-F GCCATGGAGGCCAGTGAATTCATGTCCGTAGCCATTCTAACCTCA pGADT7-MeCYP71E7-R CAGCTCGAGCTCGATGGATCCTCAATCCCATTTGTGTTTTTTAGG pGBKT7-MeCYP71E7-F GCCATGGAGGCCAGTGAATCCATGTCCGTAGCCATTCTAACCTCA pGBKT7-MeCYP71E7-R CCGCTGCAGGTCGACGGATCCTCAATCCCATTTGTGTTTTTTAG pTRV-MeCYP71E7-F CCGGAATTCCGGCCTTCAACTCGGTAGATG pTRV-MeCYP71E7-R CGCGGATCCGCGGACATCTACCGAGTTGAA qMeCYP71E7-F ATGGCTGGCTCGCCGGAATTCGTTG qMeCYP71E7-R ATTTCCTTTCACCGCTG MeEF1a-F TGAACCACCCTGGTCAGATTGGAA MeEF1a-R AACTTGGGCTCCTTCTCAAGCTCT Table 1. Primer sequence information
-
通过Phytozome14数据库检索MeCYP71E7在不同物种内的同源序列与对应氨基酸序列,借助DNAMAN软件(DNAMAN 10)进行序列比对,得到不同物种中与MeCYP71E7同源性较高的氨基酸序列,随后通过MEGA11软件,构建系统进化树。利用PlantCARE(
https://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )进行顺式作用元件的预测,采用TBtools完成结果可视化呈现。MeCYP71E7蛋白序列的保守结构域则通过NCBI在线网站(https://www.ncbi.nlm.nih.gov/ )预测。 -
取适量木薯叶片置于研钵内,倒入液氮充分研磨至粉末状态,采用CTAB法[16]提取总RNA,随之将RNA反转录成cDNA(ZR108-100T,北京,庄盟),最后将cDNA贮存于−80℃冰箱备用。
-
从在线数据库Phytozome14中获取MeCYP71E7的CDS序列,使用Primer5工具完成pCAMBIA2300-MeCYP71E7上游引物与pCAMBIA2300-MeCYP71E7下游引物的设计(表1)。以cDNA为模板,开展MeCYP71E7目标片段的PCR扩增(Mastercycler® nexus)反应,反应程序设定为:95℃预变性3 min;95℃ 30 s、52.8℃ 30 s、72℃ 2 min,32个循环;最后72℃延伸10 min。将目标条带进行胶回收纯化,随后连接pEASY-T3克隆载体,并转入DH5α感受态细胞中[17],转化完成后采用菌落PCR方法开展阳性克隆验证。筛选出电泳条带单一且亮度较高的单克隆菌落,送至测序公司完成序列测定与比对分析。将测序结果与目标基因序列完全匹配的菌株置于−40℃超低温冰箱中长期保存。后续将MeCYP71E7的完整CDS序列利用T4 DNA连接酶插入pCAMBIA2300表达载体,随之将重组载体转入DH5α感受态细胞,通过菌落PCR方法初步验证阳性克隆。挑选电泳条带单一清晰的阳性单克隆菌落进行扩大摇菌培养,之后进行质粒的提取,最终通过BamH I与Sal I双酶切验证重组表达载体的构建是否正确。
-
为了分析MeCYP71E7在木薯不同组织中的表达量,本研究利用在线数据TCOD expression(
https://shiny.datasci.danforthcenter.org/cassava_atlas/ )查找该基因在木薯不同组织中的表达水平,并用Prism8软件绘图。 -
使用Primer5软件进行MeCYP71E7定量引物的设计(表1),本研究选取延伸因子基因(Elongation factor,EF)作为内参基因,以合成的cDNA为模板,利用荧光定量PCR仪(CFX ConnectTMReal-Time)开展MeCYP71E7过表达和沉默载体检测。具体反应体系配置如下:ddH2O 6.5 μL、cDNA模板 0.2 μL、上游引物 0.4 μL、下游引物 0.4 μL,以及SYBR荧光染料7.5 μL。反应设置:95℃ 3 min;95℃ 30 s、55℃ 30 s、72℃ 20 s,45个循环。所有样品均设置3个生物学重复,通过2−ΔΔCt进行数据分析,并通过单因素方差分析法完成统计分析。
-
采用双酶切将MeCYP71E7的CDS片段插入到pCAMBIA2300载体中,成功构建35S::MeCYP71E7-GFP重组载体,随后转化至GV3101农杆菌。使用含有10 mmol·L−1 MgCl2、50 mmol·L−1 MES和150 μmol·L−1 AS的渗透液制备OD600值为0.6的农杆菌菌液,并将其置于冰上静置处理[18]。最后将农杆菌菌液注射到烟草叶片的下表皮,48 h左右取叶片,在共聚焦显微镜(LSM900,ZEISS)下,观察荧光情况。
-
为明确MeCYP71E7的抗病功能,本试验首先构建该基因的过表达与沉默木薯材料系,后续对处理后的木薯叶片开展Xam(实验室保存)接种试验。试验过程中,将重组过表达载体pCAMBIA2300-MeCYP71E7、空载pCAMBIA2300,以及pTRV1、pTRV2与沉默重组载体pTRV2-MeCYP71E7分别转入GV3101农杆菌感受态。过表达组以转入空载pCAMBIA2300的农杆菌侵染材料为空白对照(Mock-1/Mock-2),转入pCAMBIA2300-MeCYP71E7的农杆菌侵染材料为处理组(MeCYP71E7-OE-1/MeCYP71E7-OE-2),采用无菌注射器将菌液注入木薯叶片下表皮。侵染3 d后,分别采集各单株上被农杆菌侵染的叶片提取总RNA,分析目的基因的相对转录水平以验证过表达率。验证植株已过表达后,对已完成农杆菌侵染的叶片接种Xam病原菌,Xam病原菌OD600值到达0.8左右,第6天进行病斑面积及表型统计。
沉默设置浓度一致的pTRV1和pTRV2农杆菌等比例混合作为对照组(CK-1/CK-2),pTRV2-MeCYP71E7和pTRV1混合菌液作为试验组(MeCYP71E7-RNAi-1/MeCYP71E7-RNAi-2),用无菌注射器注射到木薯叶背面,侵染12 d后取上层未注射农杆菌的叶片提取RNA并反转录为cDNA,以cDNA为模板来进行RT-qPCR验证,分析沉默相对转录水平,随后,对上层未注射农杆菌的新叶片接种Xam,进行病斑面积统计。
-
取−80℃保存的AH109菌株,划线于YPDA固体平板上,置于28℃培养2~3 d;挑取平板上生长的酵母单菌落,置于5 mL的YPDA液体中,28℃摇床培养8~10 h;吸取一定量的AH109菌液(由实验室保存),置于50 mL的YPDA溶液中进行转摇,调节初始OD600值为0.2~0.3,之后置于28℃培养3~5 h,使OD600值到达0.6左右;将菌液分装在50 mL离心管中,4 000 r·min−1,低转速离心10 min,去上清;每管加人40 mL的ddH2O重悬,4 000 r·min−1,低转速离心10 min,去上清;加人2 mL的1×TE/LiAC重悬,完成酵母感受态细胞的制备。
-
ssDNA分别与pGBKT7、pGADT7、pGADT7-MeCYP71E7质粒组合各10 μL混合,将混合液置于2 mL离心管中;接着加入酵母感受态细胞0.2 mL,PEG/LiAC 0.6 mL,随后颠倒混匀。将离心管置于28℃摇床中培养1 h,随后加入70 μL DMSO,于42℃水浴环境中热激30 min;以
12000 r/min的转速离心30 s后去除上清液;用300 μL 1×TE缓冲液重悬沉淀,将其涂布在SD-Trp/-Leu和SD-Trp/-Leu/-Ade/-His培养基上,28℃进行培养。 -
将pGBKT7与pGBKT7-MeCYP71E7两种重组载体分别涂布在SD-Trp和SD-Trp/-Ade/-His固体培养基,观察其生长情况来判断pGBKT7-MeCYP71E7是否有自激活现象。实验采用4种浓度梯度稀释接种至对应培养基上。
-
使用WPS Office和SPSS 24进行数据处理和方差分析,采用单因素方差分析,当P<0.05时,代表处理间差异显著(P<0.05)。使用GraphPad Prism 8.0.1作图。
-
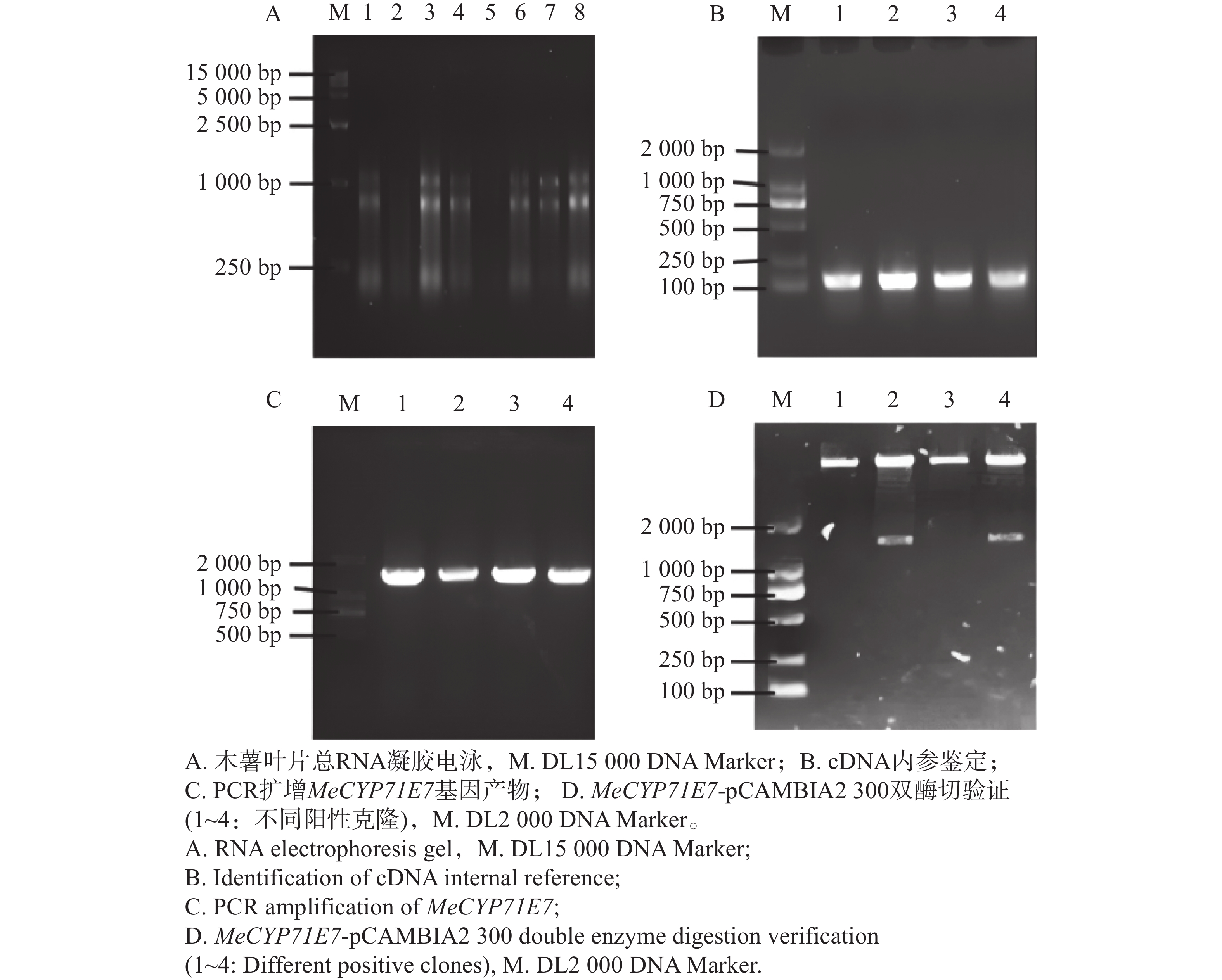
克隆MeCYP71E7基因,提取木薯叶片总RNA,经琼脂糖凝胶电泳检测其完整性,结果显示存在28S、18S和5S 3条清晰条带,说明所提取的RNA完整性较好,无明显降解(图1-A),可用于后续实验。随后将RNA反转录成cDNA,利用MeEF1a对cDNA进行PCR质量验证,扩增产物呈现单一清晰条带,说明cDNA反转录成功(图1-B)。以上述cDNA为模板进行PCR扩增,凝胶电泳在1 000 ~2 000 bp处存在特异性条带,MeCYP71E7的CDS全长为1 536 bp,扩增条带与目的基因大小一致(图1-C)。随后,对PCR产物进行胶回收,并连接到pCAMBIA2300载体,转入DH5ɑ,涂布到培养基上,接着进行菌落PCR,阳性单克隆摇菌,提取质粒,最后利用BamH I/Sal I进行酶切验证,目的条带在1 000~2 000 bp之间,即为目的条带(图1-D),并进行测序验证,证实MeCYP71E7已构建到pCAMBIA2300载体上,即MeCYP71E7过表达载体构建成功。
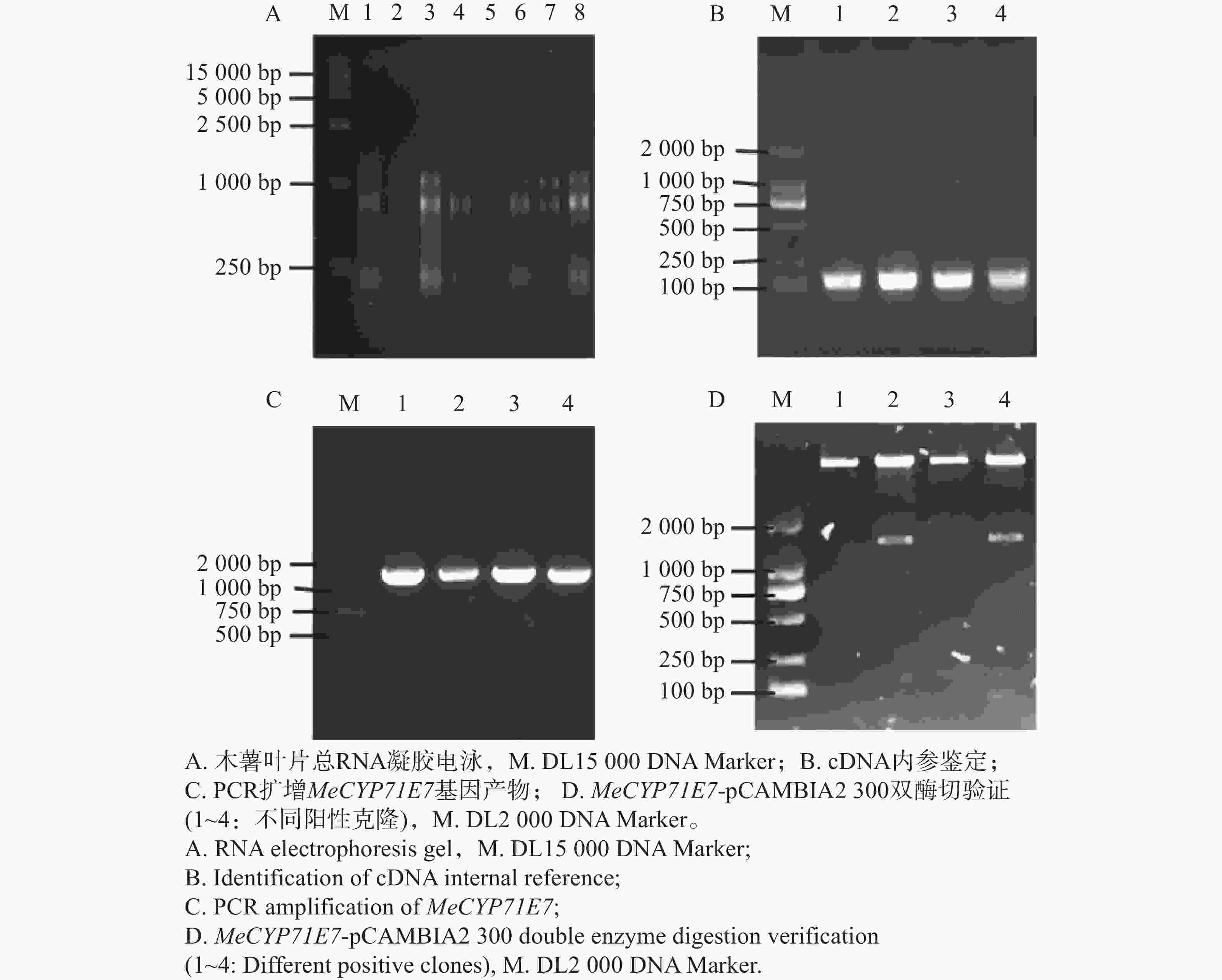
Figure 1. Amplification of the MeCYP71E7 fragment and construction the expression vector
-
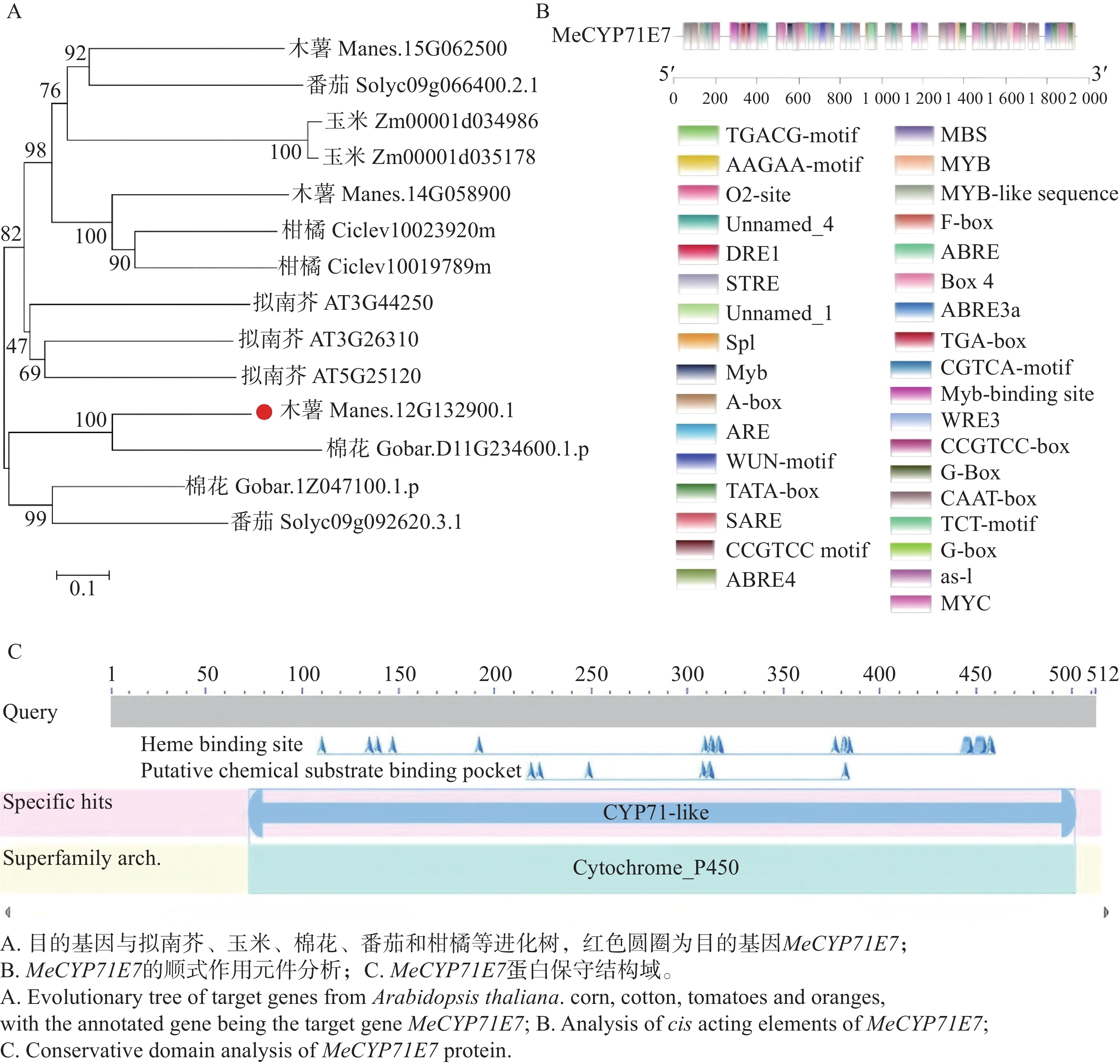
为明确MeCYP71E7基因与其他植物的进化关系。通过MEGA11软件,对木薯、玉米、棉花、番茄、柑橘、拟南芥6种植物的CYP基因和MeCYP71E7绘制进化树(图2-A)。研究发现,MeCYP71E7和棉花CYP的同源性较高。此外,对MeCYP71E7上游的序列进行顺式作用元件分析发现,MeCYP71E7启动子区域含有核心元件TATA-box,增强元件CAAT-box,激素响应元件ABRE、CGTCA等,光响应元件G-Box,还含有转录因子MYB和MYC等结合位点(图2-B)。蛋白质结构域分析发现,MeCYP71E7属于CYP450家族,属于CYP71亚家族(图2-C)。

Figure 2. Bioinformatics analysis of MeCYP71E7
-
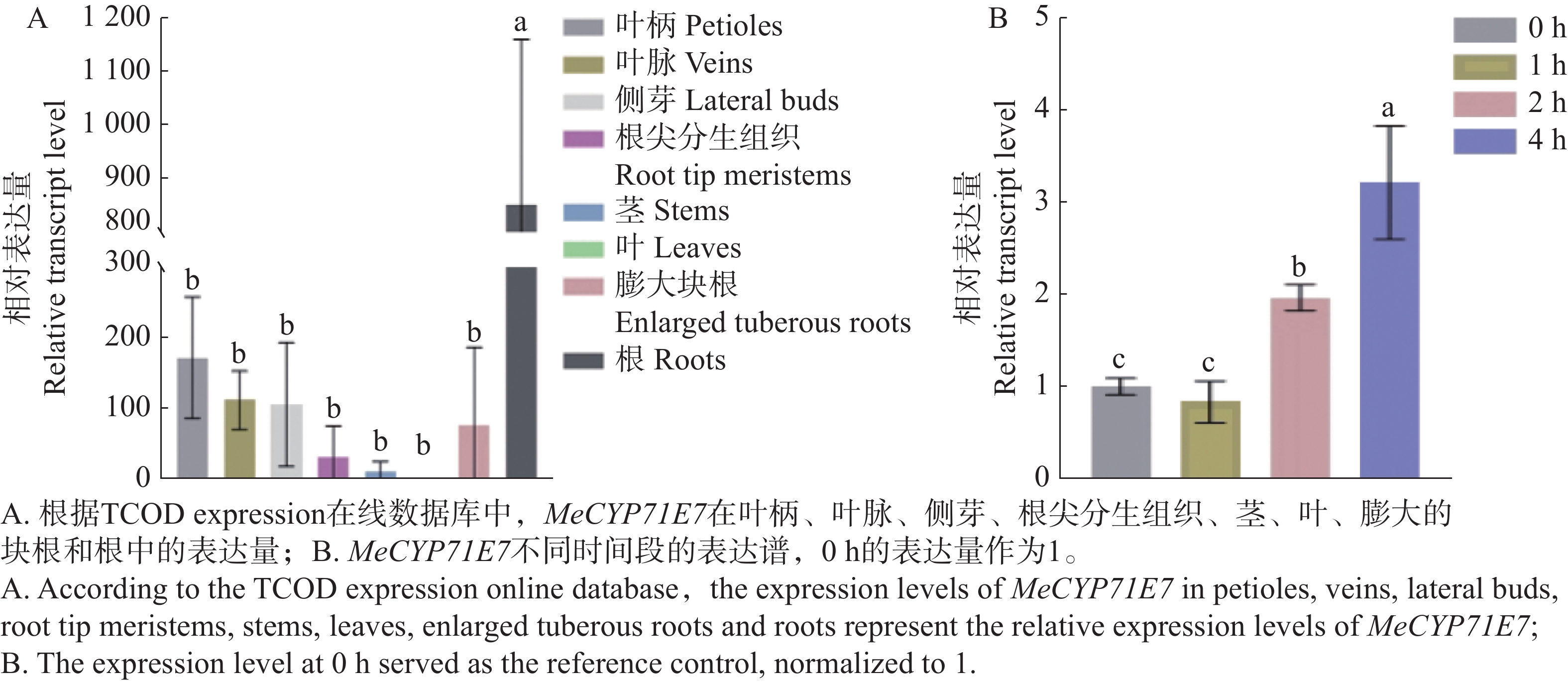
分析MeCYP71E7在木薯不同组织中的表达量,本研究利用在线数据TCOD expression查找该基因在木薯不同组织中的表达水平,并用Prism8软件绘制。MeCYP71E7在木薯的根、叶柄、叶脉、侧芽、块根、根尖分生组织、茎中均有表达(图3-A)。MeCYP71E7在木薯根中的表达量显著高于其他组织中的表达,其次为叶柄、叶脉、侧芽、块根、根尖分生组织、茎以及叶,表达量均不显著。另外,MeCYP71E7在木薯叶中的表达量虽被检测到,但表达水平最低。为进一步证实MeCYP71E7表达量是否受到细菌性枯萎病影响,通过RT-qPCR测定不同时间点的相对表达量(图3-B)。结果显示,MeCYP71E7表达量随时间延长其逐渐升高(0、1、2 、4 h)。
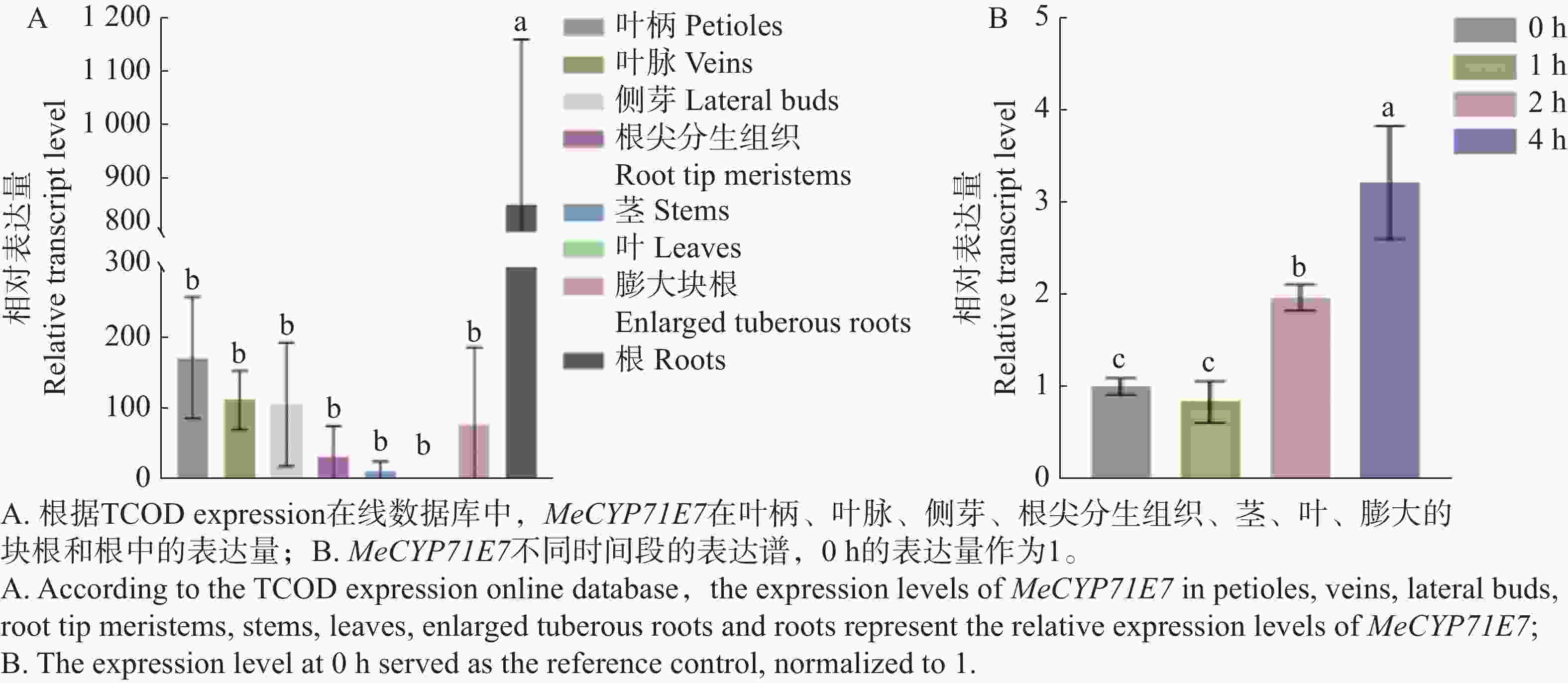
Figure 3. Expression profiling of MeCYP71E7 in different tissues of cassava
-
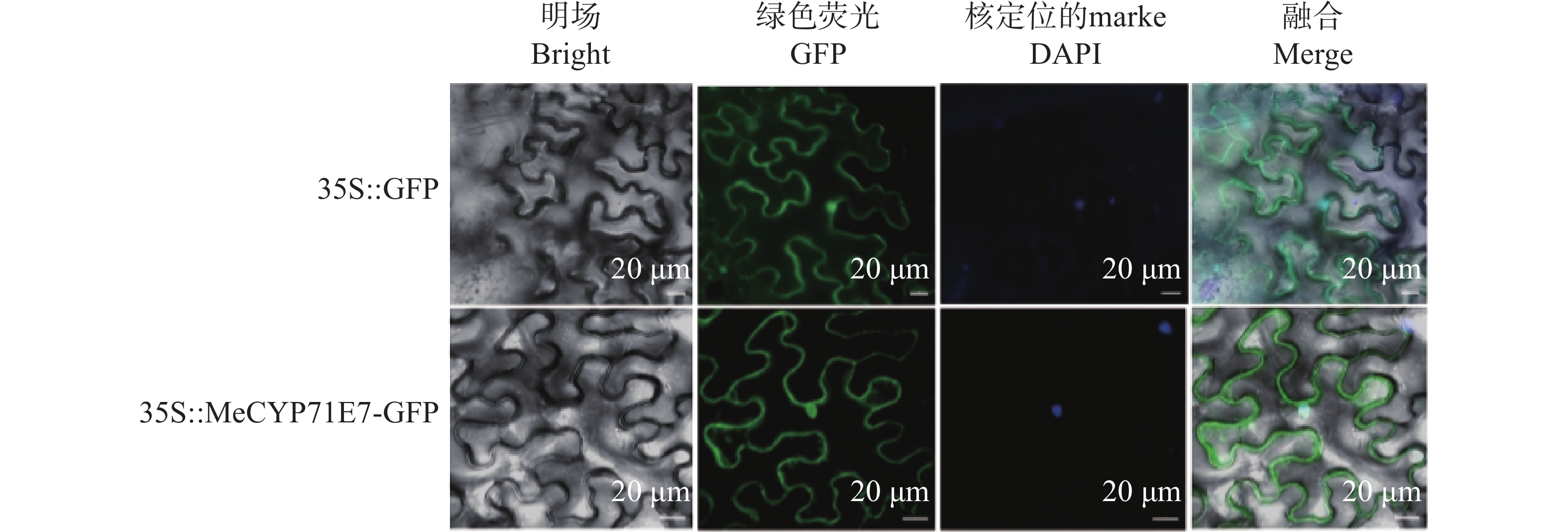
明确MeCYP71E7在木薯中的表达位置,构建35S::MeCYP71E7-GFP融合表达载体并进行瞬时表达。侵染烟草叶片背面,瞬时表达48 h后,通过共聚焦显微镜观察叶片发现,在35S启动子的驱动下,绿色荧光蛋白可以与MeCYP71E7融合表达,其表皮细胞的细胞核、质膜上存在绿色荧光。为进一步验证定位的准确性,将观察到绿色荧光的叶片用细胞核染色试剂DAPI(4',6-diamidino-2-phenylindole,DAPI)染色2.5 h,共聚焦荧光显微镜下看到DAPI所发出的蓝色荧光所显示的细胞核位置与MeCYP71E7-GFP的绿色荧光相一致,叠加后位置重合(图4)。结果表明,MeCYP71E7定位于细胞核、细胞膜。
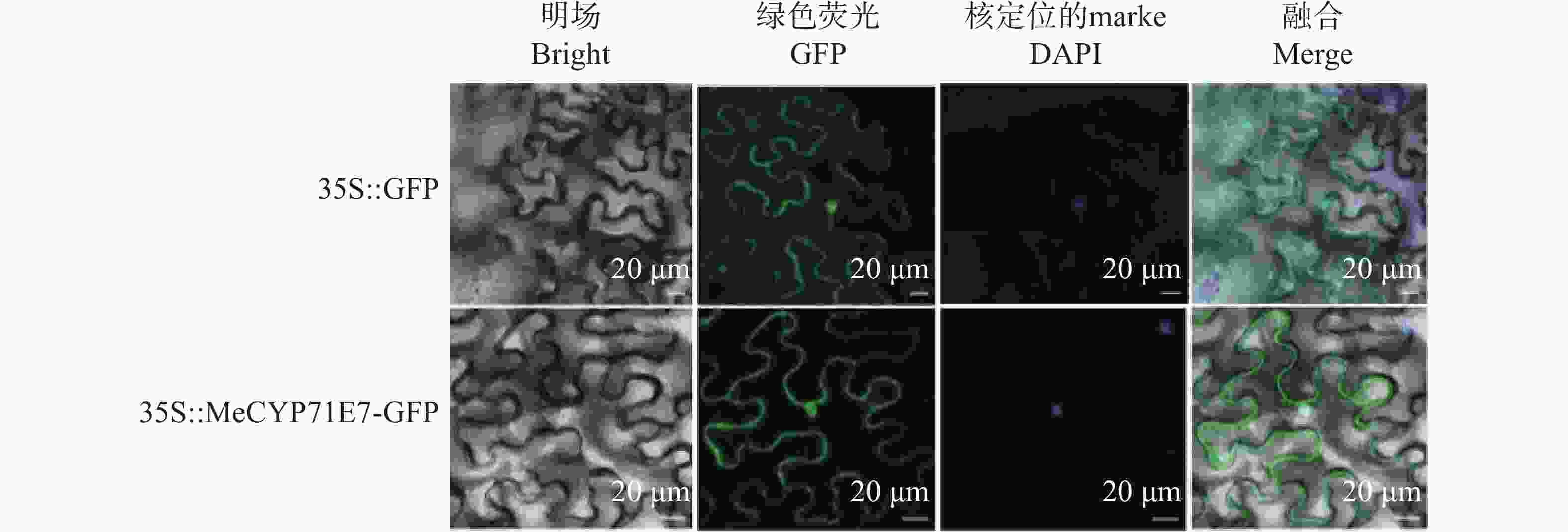
Figure 4. Subcellular localization of MeCYP71E7
-
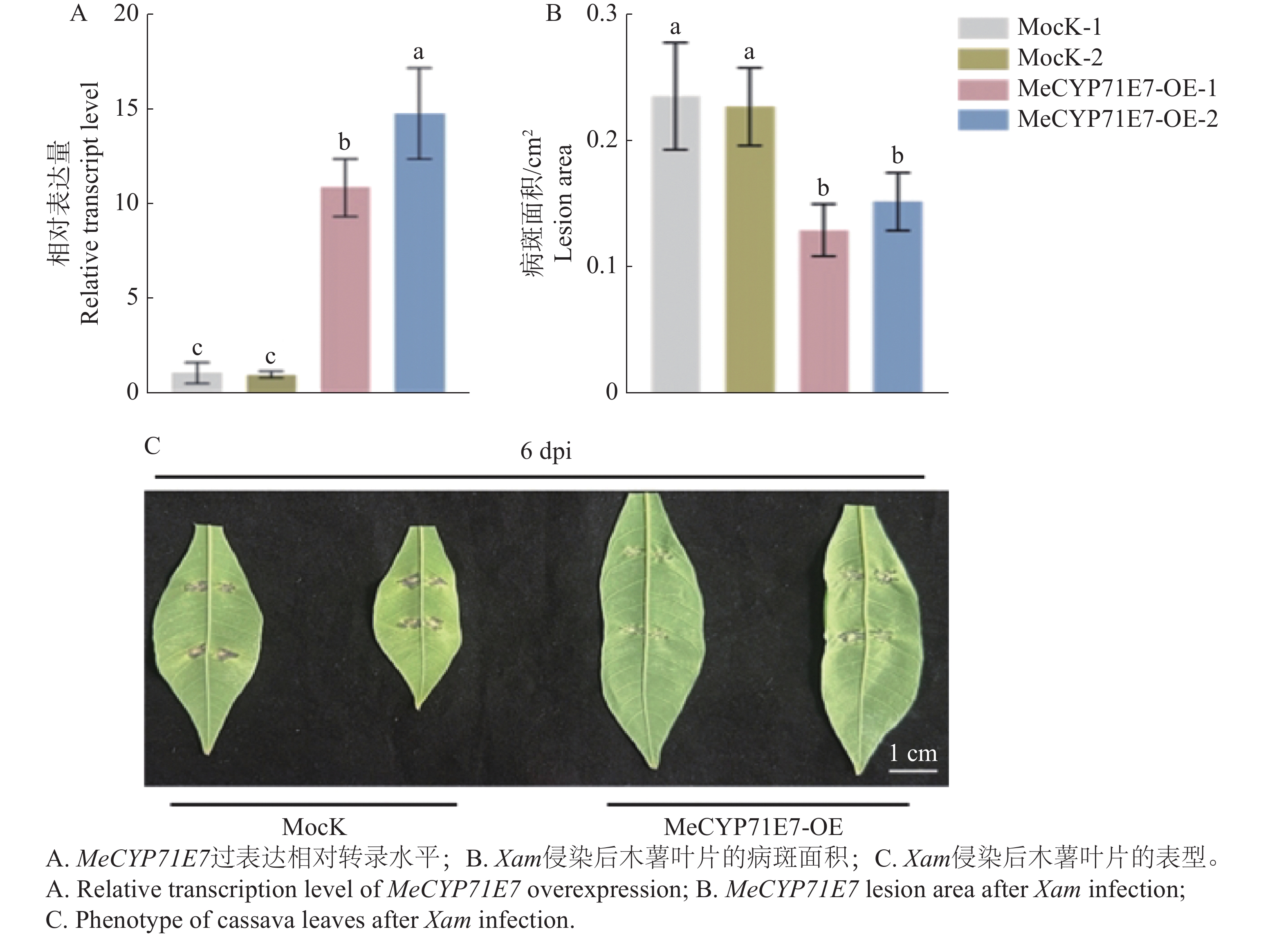
探究MeCYP71E7对Xam侵染后抗病的影响,本研究构建MeCYP71E7过表达株系,并对木薯叶片接种Xam。过表达MeCYP71E7植株中的MeCYP71E7的表达量显著高于对照组(5-A),说明木薯MeCYP71E7过表达植株构建成功。同时对病斑面积进行了统计,实验显示,与对照相比,MeCYP71E7病斑面积显著降低(图5-B),MeCYP71E7发病情况明显轻于对照组(图5-C),推测MeCYP71E7基因参与调控木薯的抗病性。
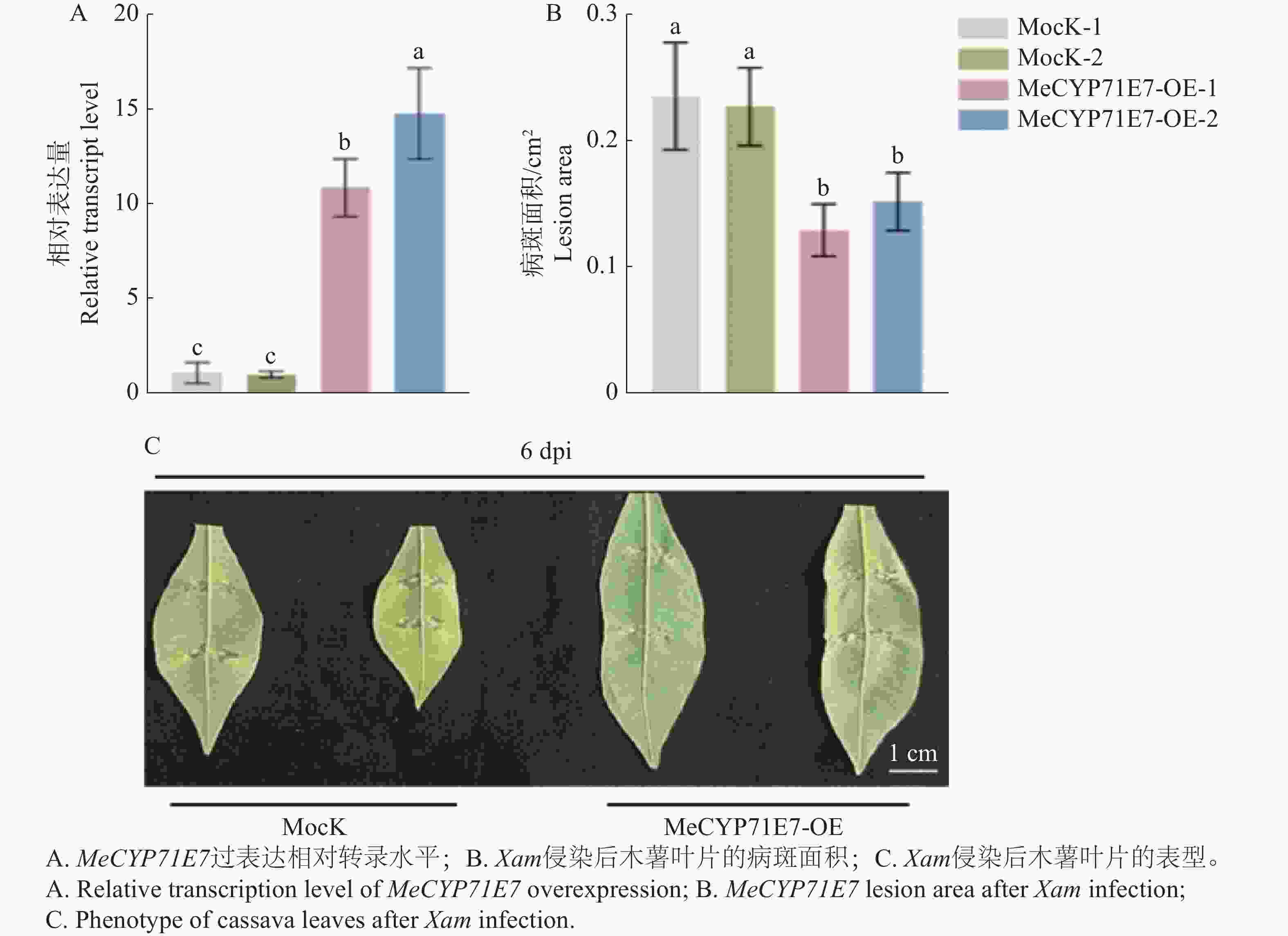
Figure 5. The effect of MeCYP71E7 overexpression on cassava disease resistance
-
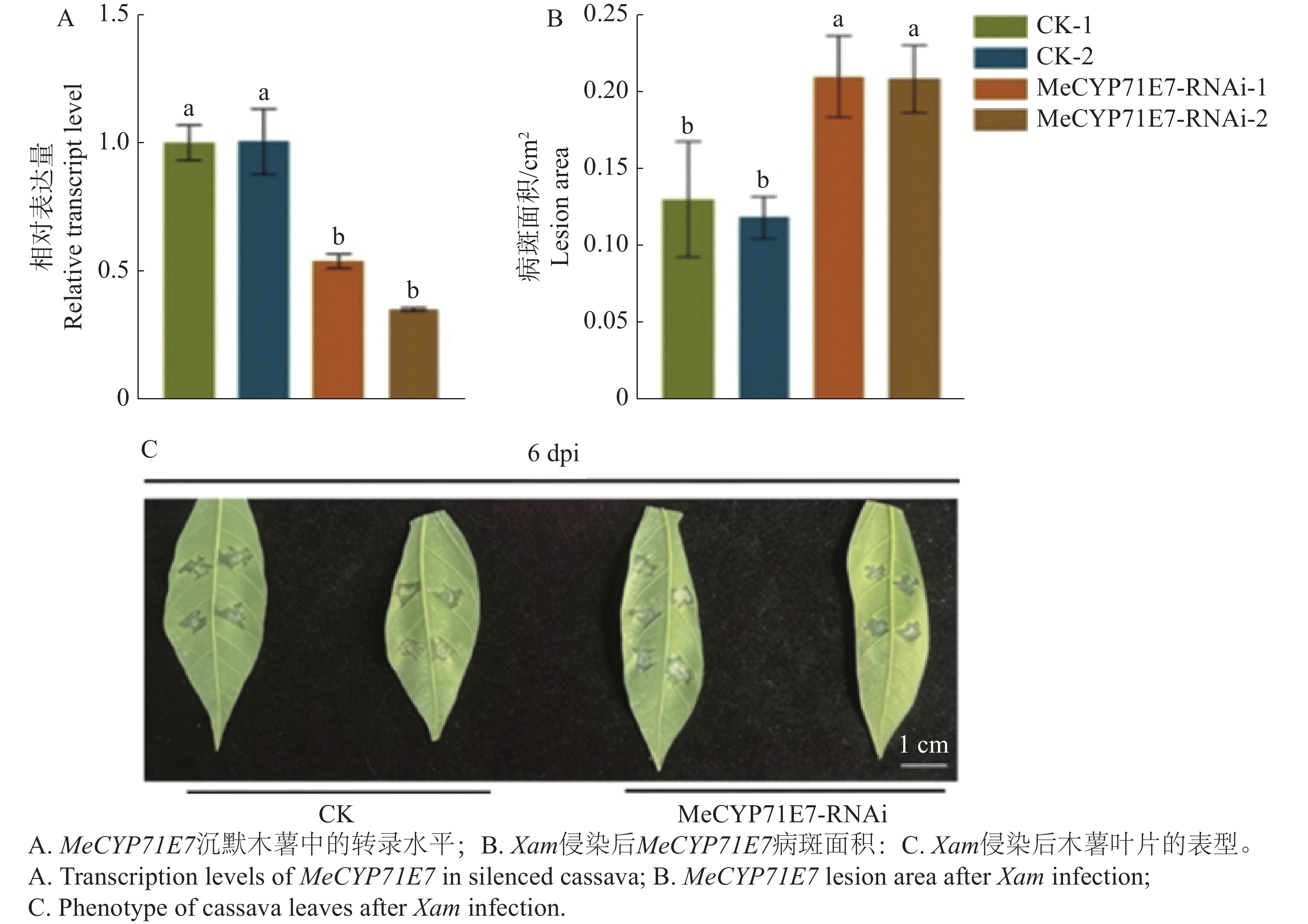
进一步确定MeCYP71E7抗病性,本研究建立了MeCYP71E7沉默植株体系。结果显示,MeCYP71E7-RNAi-1和MeCYP71E7-RNAi-2木薯植株中的表达量降低,表明MeCYP71E7沉默株系构建成功(图6-A)。进一步分析叶片的病斑面积,实验显示,MeCYP71E7沉默病斑面积扩大(图6-B),发病情况明显重于对照组(图6-C)。可见,MeCYP71E7正调控木薯抗病性。
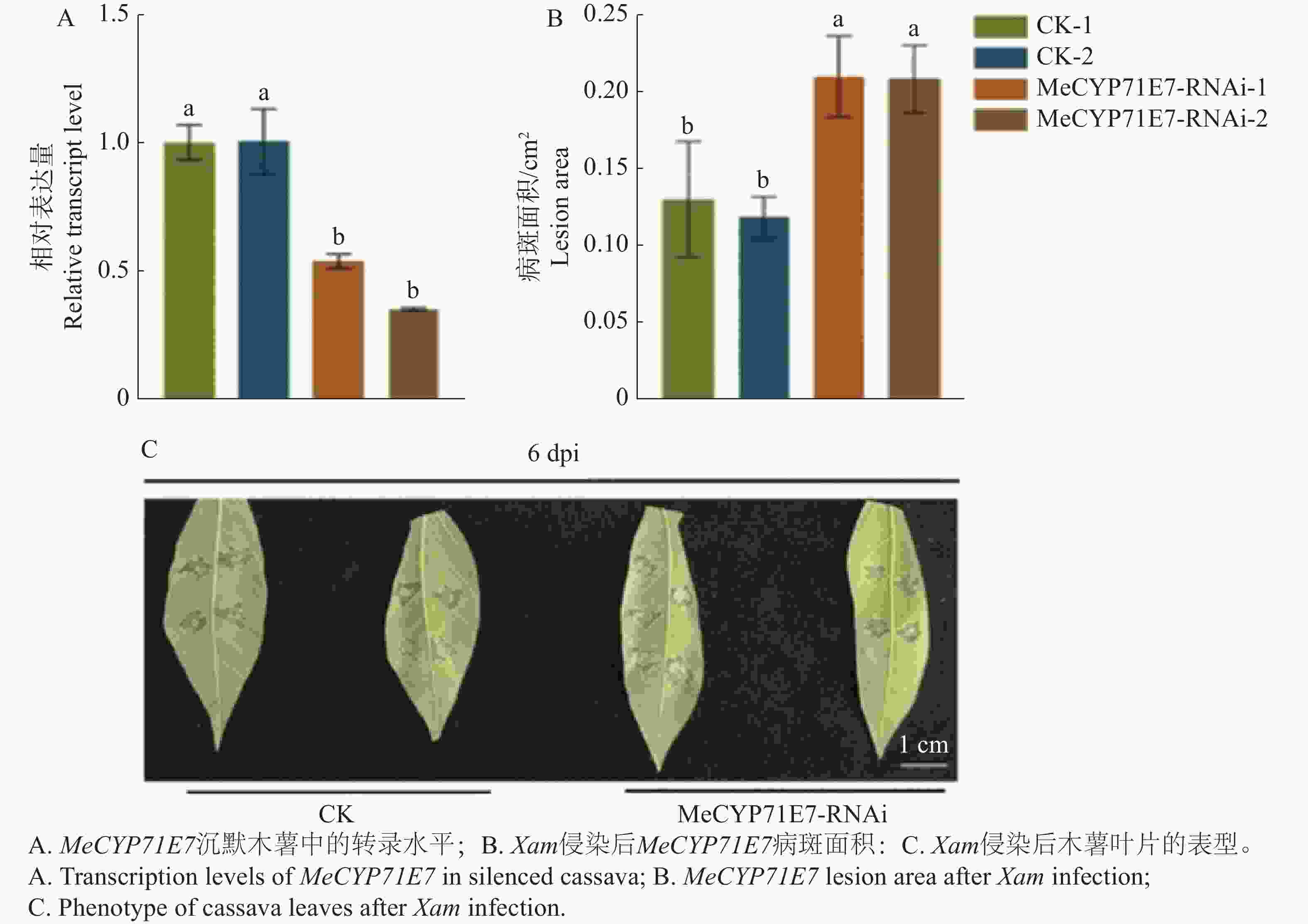
Figure 6. The effect of MeCYP71E7 silenced on cassava disease resistance
-
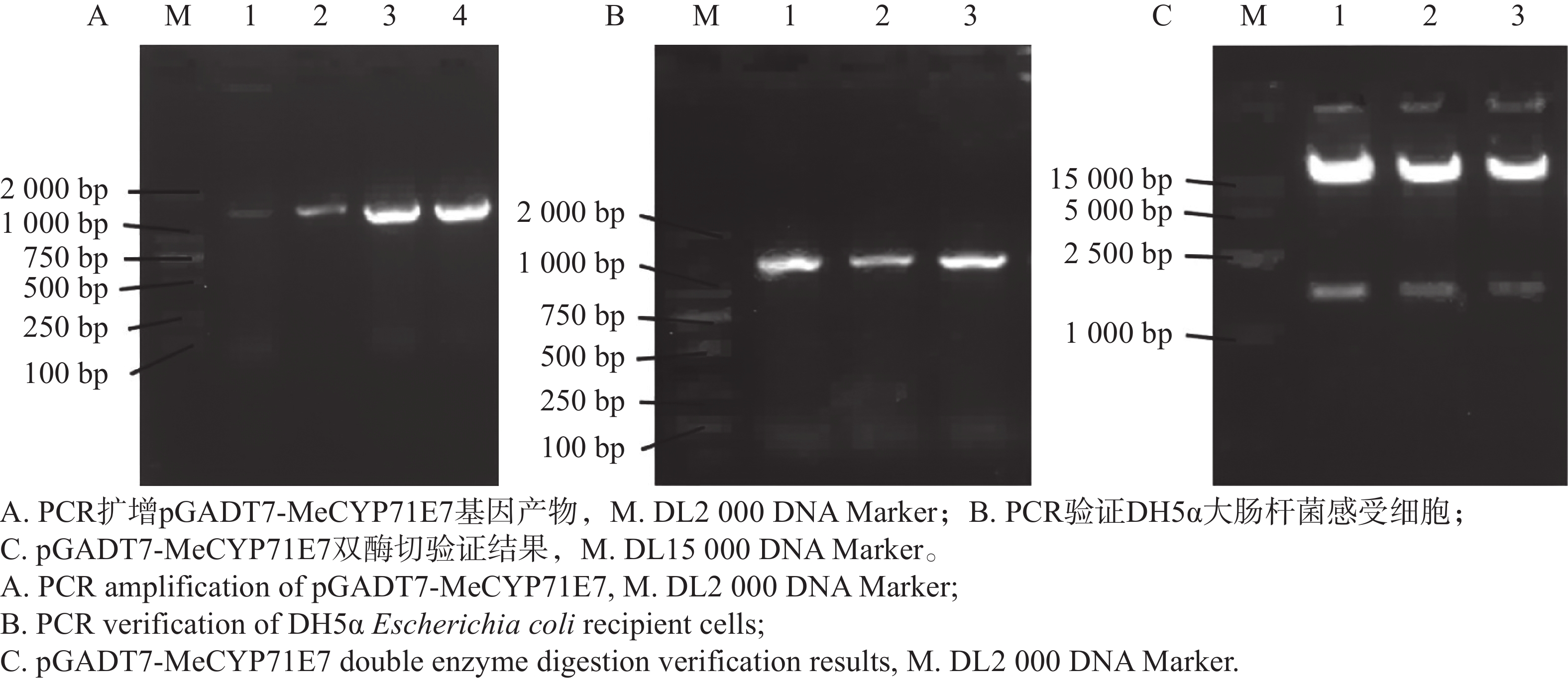
进一步解析MeCYP71E7调控木薯抗病的分子机制,我们构建MeCYP71E7的pGADT7载体。以木薯cDNA为模板,用引物pGADT7-MeCYP71E7进行PCR扩增,凝胶电泳检测到片段大小约1 536 bp与目的基因实际长度一致(图7-A)。利用同源重组技术将目的片段连接到EcoR I/BamH I双酶切pGADT7载体上。连接产物转化到DH5α大肠杆菌感受细胞中,挑选单菌落作为模板并PCR验证。结果显示,扩增片段同目的基因克隆长度相同(图7-B)。挑选5个阳性克隆,对其质粒酶切验证(图7-C)并测序检验。保留测序结果与目的基因一致的克隆用于后续酵母双杂实验。

Figure 7. Construction of pGADT7-MeCYP71E7 gene vector
-
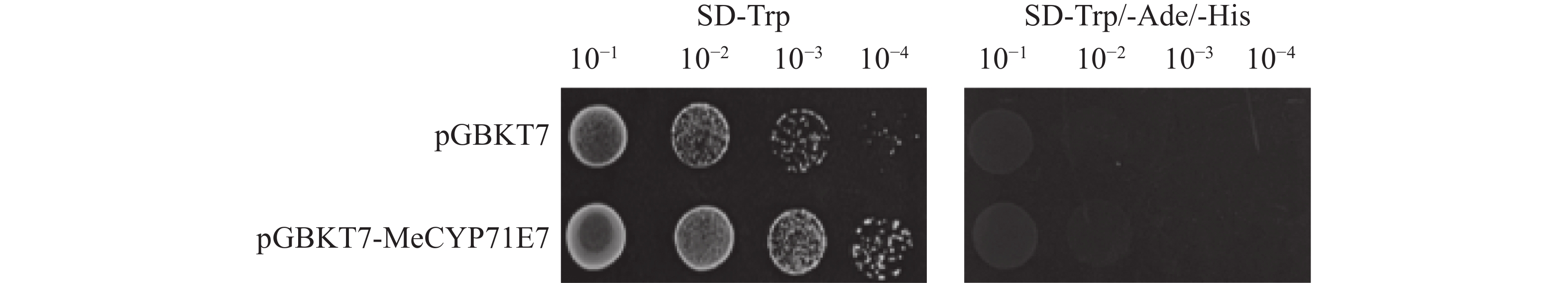
验证MeCYP71E7是否存在自激活。本研究将重组质粒pGBKT7-MeCYP71E7和空载质粒pGBKT7分别转入AH109,同时以空载pGBKT7作为对照。随后将稀释为4个梯度的酵母菌液分别点涂于SD-Trp(一缺)和SD-Trp/-Ade/-His(三缺)固体培养基上,并观察酵母生长状况。实验结果表明,在在一缺培养基上,两组酵母能够正常生长,且酵母数量随菌液稀释倍数的增加而逐渐减少。而在三缺培养基上,两组处理均未出现酵母生长(图8)。可见,MeCYP71E7不存在自激活活性。
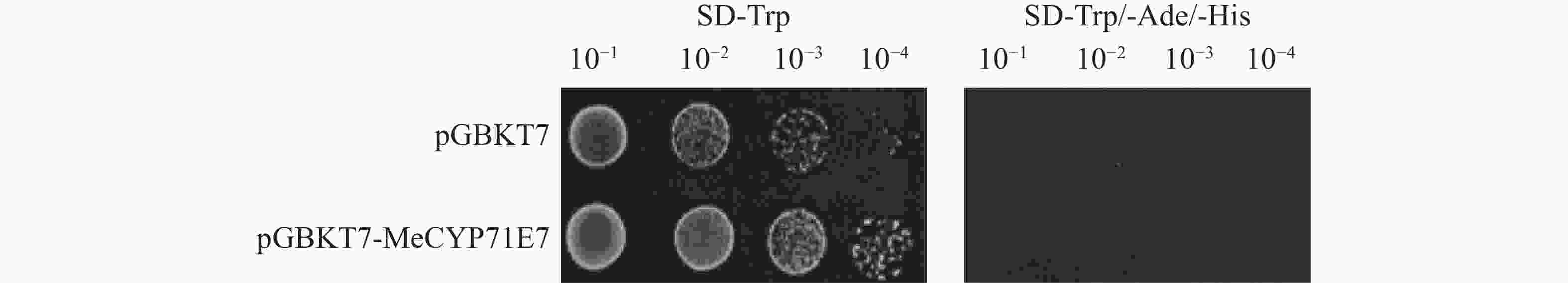
Figure 8. The self-activation validation of pGBKT7-MeCYP71E7
-
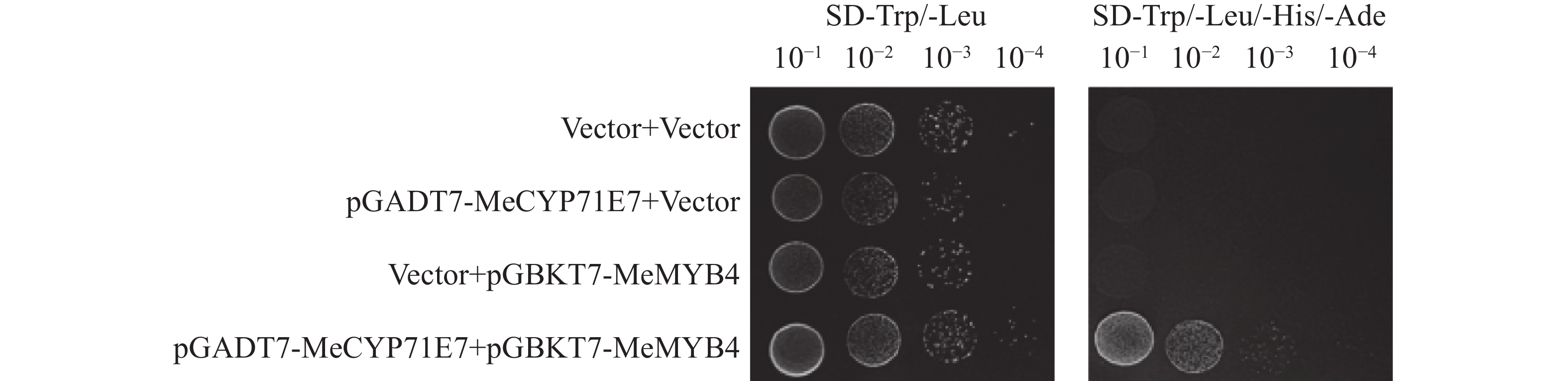
进一步探究MeCYP71E7调控木薯抗病反应的分子机制,本研究实验室前期使用酵母双杂交进行筛库,初步筛选到MeMYB4,通过点对点验证MeCYP71E7与MeMYB4互作。酵母双杂交实验结果表明,pGADT7-MeCYP71E7和pGBKT7-MeMYB4共转化AH109,在二缺(SD-Trp/-Leu)和四缺(SD-Trp/-Leu/-His/-Ade)的培养基上均可正常生长,并且随着菌液浓度(10−1、10−2、10−3、10−4)稀释,生长的菌落呈现梯度减少趋势,而其他3组酵母菌在二缺培养基能正常生长,四缺培养基无法生长(图9)。结果表明MeCYP71E7与MeMYB4之间是互作的。
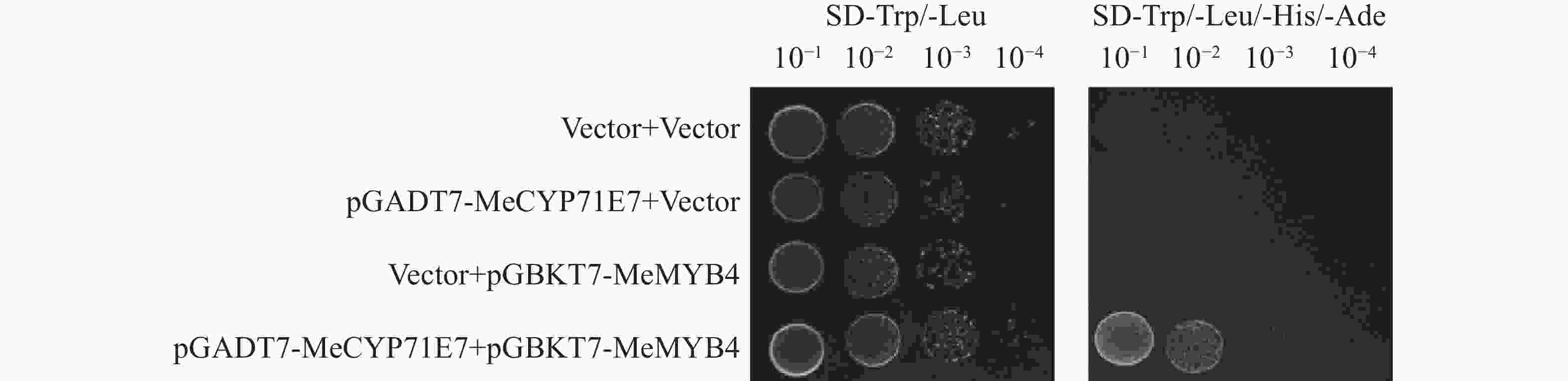
Figure 9. The Validation of the interaction between MeCYP71E7 and MeMYB4
-
细胞色素 P450(CYP450)是一类以血红素为辅基的末端加氧酶,因在 450 nm 波长下存在特征吸收峰而得名 [19]。CYP71 家族隶属于 CYP450 超家族的 CYP71 簇,该家族成员数量最为丰富,在植物次生代谢合成通路中发挥关键功能 [20−21]。整体而言,CYP450 超家族是植物代谢网络的核心功能蛋白 [22]。在木薯中,CYP71E7 属于肟代谢关键酶,直接参与氰苷生物合成;本研究围绕木薯 MeCYP71E7 基因,系统解析其调控木薯细菌性枯萎病抗性的分子功能。
本研究成功从木薯中克隆得到 CYP450 超家族基因 MeCYP71E7,蛋白结构域分析结果证实该基因归属于 CYP450 超家族 CYP71 亚家族。CYP450 基因家族在各类植物中均高度保守,保守结构域可将血红素稳定锚定在蛋白活性中心,维持蛋白核心构象完整 [23]。对 MeCYP71E7 启动子序列开展顺式作用元件预测分析,结果显示,该启动子包含 TATA-box 核心启动子元件、CAAT-box 增强子元件以及多种植物激素响应顺式元件,预示 MeCYP71E7 的转录表达受多条激素信号通路协同调控。烟草 CYP82E4 启动子同样富集 TATA-box、CAAT-box 及各类激素响应元件,与本研究结果一致 [24]。组织表达模式检测表明,MeCYP71E7 在木薯不同组织器官中表达水平差异显著,其中根部表达量最高,叶柄次之。已有研究报道,大豆(Glycine max(L.)Merr.)GmCYP82D26、黄瓜(Cucumis sativus L.)CsCYP82D47 与 CsCYP82D102 均在根系优势表达,与本研究组织表达特征相符 [23,25]。亚细胞定位结果显示, MeCYP71E7 定位于细胞核与细胞膜;与之不同的是,大豆 GmCYP82D26 瞬时表达于烟草表皮细胞时定位于内质网 [25],推测该类 CYP71 家族蛋白的亚细胞分布存在物种特异性,功能分化或与此相关。
CYP450 蛋白可介导多条次生代谢通路合成防御类活性物质,通过积累毒性代谢产物提升植物对昆虫、病原菌等生物胁迫的耐受能力 [26]。本研究抗病表型鉴定结果显示,过表达 MeCYP71E7 能够显著缩小木薯叶片细菌性枯萎病病斑面积,沉默 MeCYP71E7 则会加剧病斑扩展,证明 MeCYP71E7 对木薯细菌性枯萎病抗性具有正向调控作用。现有诸多研究也证实 CYP71 亚家族成员为植物抗病正调控因子:水稻 CYP716A16 过表达可增强植株对立枯丝核菌、稻黄单胞菌的抗性 [27];拟南芥 CYP82C2、CYP71A12/13 催化含氰吲哚防御物质 4-OH-ICN 合成,相关基因突变体对丁香假单胞菌敏感性大幅上升,明确了 CYP450 介导氰基代谢产物在植物免疫中的核心地位 [28];水稻 CYP71Z2 稳定过表达可长期增强植株抗细菌性枯萎病能力 [11]。
为进一步阐明 MeCYP71E7 介导抗病性的分子机制,本研究筛选其互作靶蛋白。自激活检测证明 MeCYP71E7 诱饵载体无自主转录激活活性,与水稻 OsCYP22 诱饵载体特性一致 [29];酵母双杂交试验进一步验证 MeCYP71E7 可与转录因子 MeMYB4 发生蛋白互作。已有研究表明,遭受生物胁迫时,植物可激活 CYP450 依赖的次生代谢通路,合成具备抑菌、抗虫活性的代谢物以抵御病虫害 [30];梨(Pyrus spp)CYP714C2 与 GA2ox1 互作共同调控果实抗病能力 [31];拟南芥 AtCYP71 结合组蛋白 H3,参与调控植物器官建成进程 [32];梨 PpyMYB144 可转录激活细胞色素 P450 基因 PpyCYP86B1,调控果皮褐变过程 [33];番茄(Solanum lycopersicum L.)CYP94C1 敲除后内源防御激素积累,植株病原菌抗性显著提升 [34];棉花( Gossypium hirsutum L.)CYP450 通路通过重塑黄酮代谢通量抑制枯萎病发病进程 [35]。结合本实验结果推测,MeCYP71E7 与 MeMYB4 形成蛋白复合物,协同激活下游抗病应答通路,进而正向调控木薯细菌性枯萎病抗性。
综上,本研究明确 MeCYP71E7 是木薯免疫应答的正向调控基因。但 MeMYB4 与 MeCYP71E7 协同介导抗病的完整分子通路仍较为复杂,相关调控环节有待进一步验证。后续可深入解析 MeCYP71E7 调控木薯抗细菌性枯萎病的完整分子网络,为木薯抗病分子育种、稳产增产提供理论支撑。
-
MeCYP71E7为CYP450家族,属于CYP71亚家族,并且其启动子存在多种顺式作用元件。MeCYP71E7定位于细胞核等,且在不同部位表达量不同。进一步通过过表达和基因沉默证实,MeCYP71E7正调控对木薯细菌性枯萎病抗性。
Cloning and functional analysis of MeCYP71E7 gene in cassava
DOI: 10.15886/j.cnki.rdswxb.20260074
- Received Date: 2026-05-12
- Accepted Date: 2026-06-23
- Rev Recd Date: 2026-06-17
-
Key words:
- Cassava /
- MeCYP71E7 /
- disease resistance /
- gene cloning /
- functional analysis
Abstract: The cloning, expression pattern and functional analysis of the cassava MeCYP71E7 gene were conducted, which provided a scientific basis for exploring the disease resistance of cassava and the disease resistance mechanism mediated by its interacting proteins. A bioinformatics analysis of the MeCYP71E7 gene was conducted to explore its phylogenetic relationship and analyze the conserved structure of its protein. Furthermore, the localization of the gene was determined through transient expression. Overexpression and gene-silenced plants were identified by quantitative PCR. Finally, interacting proteins were screened through yeast two-hybrid. The cDNA of cassava leaves was used as the template to successfully clone the MeCYP71E7 gene. The sequencing results were consistent with the sequence on the Phytozome database. MeCYP71E7 contained the CYP71-Like conserved domain and belonged to the CYP450 family and the CYP71 subfamily. Meanwhile, the promoter region of MeCYP71E7 contained core elements such as TATA-box, enhancer elements such as CAAT-box, and hormone response elements such as ABRE and CGTCA. In addition, the expression levels of MeCYP71E7 in different parts were analyzed, and it was found that the expression level of MeCYP71E7 was higher in roots, followed by petioles, and the lowest in leaves. The coding sequence of MeCYP71E7 was inserted into the pCAMBIA2300 vector via double restriction enzyme digestion to generate the 35S::MeCYP71E7-GFP fusion plasmid. The subcellular localization analysis indicates that the protein MeCYP71E7 is located in the nucleus and other parts of the cell. Overexpression and silencing plant systems were constructed. The number of bacteria and the area of lesions on the leaves of cassava overexpressing MeCYP71E7 decreased, while the results were opposite in the silenced cassava lines. Yeast two-hybrid experiments proved that the protein MeCYP71E7 had no self-activation and interacted with MeMYB4 ex vivo. The results indicate that MeCYP71E7 positively regulates the disease resistance of cassava, which provides a theoretical basis for further research on the disease-resistant molecular mechanism of MeCYP71E7.
| Citation: | Liang Mengwei, Liu Guoyin. Cloning and functional analysis of MeCYP71E7 gene in cassava[J]. Journal of Tropical Biology. doi: 10.15886/j.cnki.rdswxb.20260074

|
 DownLoad:
DownLoad: