-
根结线虫(Meloidogyne spp.)是全球最具破坏性的植物寄生线虫类群之一,可侵染超过 3 000 种寄主植物,每年造成的农业经济损失估计超过1 000 亿美元[1-2]。尽管化学杀线剂在根结线虫防控中仍发挥重要作用,但其生态毒性高、环境持久性强,并对非靶标生物具有显著危害,且随着环境法规日益严格,其应用正持续受到限制[3-4]。因此,开发来源安全、低毒高效且具有特异作用机制的天然产物杀线剂已成为可持续农业的重要方向。在众多天然产物中,生物碱类分子由于结构多样、靶标特异性强,尤其常作用于高度保守的神经受体或代谢关键酶,被认为具有成为杀线剂先导结构的潜力[5-6]。这些证据都表明,以植物来源的生物碱为基础发展新型杀线剂,是探索安全、高效、环境友好防控策略的关键途径。
Waltherione A 是1种具有独特结构的喹诺酮类生物碱,其特征性的氧杂双环[3.2.1]辛烷融合骨架最早由 Hoelzel 等[7]从 Waltheria douradinha 的根部分离得到。随后,该化合物陆续从 Melochia chamaedrys 和 Melochia odorata 等多种植物中被分离鉴定[8-9]。更进一步的研究表明,Waltherione A 对多种疾病相关生物具有生物活性,如抑制癌细胞增殖、展现抗 HIV 活性[10],并对锥虫等寄生性原生动物表现出强烈抑制作用[11-13]。尤其值得关注的是,Waltherione A 对南方根结线虫(Meloidogyne incognita)、北方根结线虫(Meloidogyne hapla)、花生根结线虫(Meloidogyne arenaria)乃至松材线虫(Bursaphelenchus xylophilus)均表现出极强的杀线活性,其 LC50 值仅为 0.09~3.54 mg·L−1[14],远低于多数已登记的杀线剂,显示其作为新型天然来源杀线剂的巨大应用潜力。然而,尽管 Waltherione A 的杀线活性已得到充分验证,但其分子作用机制至今尚未被系统阐明。
秀丽隐杆线虫(Caenorhabditis elegans)因其基因组完全注释、体内透明、生命周期短以及与植物寄生线虫高度保守的神经、发育与代谢通路,是解析天然产物毒性机制与发现分子靶标的核心模式生物[15-17]。其神经系统结构明确,嗅觉神经元(如 AWA、AWC和ASER)及 cGMP 信号通路的功能与根结线虫具有显著同源性,为研究杀线剂的神经作用机制提供重要支撑[18-19]。此外,两者在凋亡、溶酶体、能量代谢及多种解毒酶系统上同样具有高度保守性[20-21],确保以 C. elegans 获得的作用机制可用于推断植物寄生线虫的潜在靶标。值得注意的是,已有多项研究成功利用 C. elegans 揭示杀线剂的作用模式。例如阿维菌素靶向谷氨酸门控氯离子通道导致线虫麻痹[22],选择素类天然产物需通过 P450 酶生物活化产生毒性[23],而三氟吡啶胺通过作用于线粒体 SDH 复合物并扰乱解毒酶系统导致线虫死亡[24]。此外,纳米颗粒、细菌毒素 PtxA以及挥发性化合物 2−十一酮的杀线机制也通过 C. elegans 模型被成功解析[25-27]。
C. elegans 成熟的 RNAi 反向遗传系统是探索药物靶标不可替代的工具[28-31]。该体系支持全基因组范围内的高通量功能筛选,使研究者得以在短时间内定位与药物敏感性高度相关的关键基因。例如,Shin 等 [32]利用 C. elegans 开展生殖系毒物筛查,识别了多种能导致胚胎致死的化合物;Saur 等 [33]利用全基因组 RNAi 筛选鉴定了精神药物的关键靶点 ACR-7。此外,多项研究利用该模型深入解析化学暴露对产卵量、孵化率等关键生殖终点的影响[34]。
因此,本研究首先对以 EMS 诱变构建Waltherione A抗性秀丽隐杆线虫突变群体为起点,通过正向遗传学筛选获得稳定抗性的Waltherione A突变体A1进行基础表型研究,并结合SNP mapping和全基因组重测序筛选潜在靶基因。随后利用RNAi验证靶标基因进一步筛选出候选靶标基因,而后利用分子对接模拟Waltherione A 与候选靶标基因的靶蛋白的相互作用模式,对Waltherione A的作用机制进行初探。本研究共同构建了Waltherione A在C. elegans中的作用网络框架,旨在为该天然产物作为新型杀线剂的靶标发现和理性结构优化提供依据。
-
化学品:Waltherione A(海南大学孙然锋课题组)。菌株:N2 Bristol、CB4856 Hawaiian、Escherichia Coli OP50(来源于CGC);Mutant-A1(A1,来源于广西科学院);感受态细胞E.coli DH5α和E.coli HT115、L4440质粒载体(北京擎科生物科技股份有限公司)。试剂及试剂盒:2×Taq Plus Master (Dye Plus)(南京诺唯赞生物科技股份有限Waltherione A公司)、QuickCut™ Dra Ⅰ(Aha Ⅲ)(TaKaRa),BeyoFast™XbaⅠ、BeyoFast™SacⅡ(上海碧云天生物技术有限公司),Solarbio T1410 T4 DNA连接酶(北京索莱宝科技有限公司),Tsing Zol Total RNA Extraction Reagent TsingZol、HiScript III RT SuperMix for qPCR(+ gDNA wiper)、ChamQ Universal SYBR qPCR Master Mix(北京擎科生物科技股份有限公司)。
-
将op50菌株(−80 ℃保存)划线至LB固体培养基中,37 ℃倒置培养12~14 h ,后挑取平板中生长出的单菌落至100 mL LB液体培养基中,在摇床中(200 r·min−1 ,37 ℃)培养至OD600=0.2后,取出放入4 ℃冰箱备用。
-
取适量OP50菌液加入至NGM平板中,均匀涂布于平板表面,室温过夜培养,制成NGM食物板,后切下含有线虫的NGM块转移到新的NGM食物板中,传代培养,培养温度20 ℃,其中需避免线虫污染或饥饿。
-
本实验采用限时产卵法和过滤法进行同步化。
1)限时产卵法:使用Pick挑起足够的母虫放在新的NGM食物板中,产卵2 h后将母虫烧死,放入20 ℃培养箱中,培养48 h后得到L4阶段的线虫群体。
2)过滤法:将秀丽隐杆线虫用M9冲洗下来,后使用 Millipore(11μm)滤膜进行过滤,过滤得到L1 时期的线虫,在20℃的培养箱中培养48 h 后,得到L4 阶段的线虫群体。
-
将同步化后的秀丽隐杆线虫用无菌的M9缓冲液洗净后,分别置于96孔板中进行处理。处理组包括:以M9缓冲液为溶剂的3 mg·L−1 Waltherione A处理组和6 mg·L−1 Waltherione A处理组。同时,设置空白对照组(仅含纯M9缓冲液)。所有处理均在20 °暗环境中暴露24 h。每组3个重复。
-
使用2、5、8、10 mg·L−1 的Waltherione A溶液(以M9缓冲液作为溶剂)对秀丽隐杆线虫N2进行室内杀线虫活性测定,在96孔板100 μL体系中挑取50~100条的L1阶段线虫,分别于24、48 、72 h 3个阶段统计线虫死亡率。
-
同步化方法如1.2.3,施药暴露方法如1.2.4。将暴露后的线虫用M9缓存液清洗2~3遍,至药物洗净,后放入NGM空白板中,使用Pick挑取活性较好的15条L4时期雌雄同体的线虫,接种于中央处有1滴OP50菌液的NGM平板上,单虫单板,置于20 ℃恒温培养箱中。每12 h将线虫转移到新鲜制备的OP50 1滴板中,对原培养平板在体式显微镜下进行产卵量的精确记录。对每条线虫进行持续观察,记录其产卵情况,直至观察到产卵停止(即连续多个周期内无新增虫卵产生)。最终,汇总各周期(12 h为1个周期)的产卵数据,计算每条线虫整个生命周期的总产卵量。
-
1.2.6.1中的挑走母虫的含卵原平板继续于20 ℃恒温培养箱养24 h,以确保卵完成孵化。随后,在体式显微镜下对平板中的未孵化卵(含死卵及未受精卵)数量进行计数并记录。线虫的总产卵量定义为整个生殖期内产生的所有卵(包括后续产出的卵)。孵化卵数由总产卵量减去累计未孵化卵数获得,后代存活率则通过以下公式计算:
-
待1.2.6.1中的挑走母虫的原平板培养72 h后,统计雄虫数量。
-
同步化方法如1.2.3,获得L1线虫群体,施药暴露方法如1.2.4,暴露后将线虫吸出,使用M 9缓冲液清洗3遍,等量加入至NGM食物板中,每板包含约100条线虫,后统计0 h-L1、48h-L4、72h-幼成虫3个阶段时刻中线虫长到对应时期的数量,各组浓度设置3个重复,实验重复3次。
-
同步化方法如1.2.3,同步培养至L4时期,施药暴露方法如1.2.4。将暴露后的线虫用M9缓冲液清洗3遍后吸出放至NGM空白板中,用1 mmol·L−1的左旋咪唑固定线虫,置于2 %(m/m)的琼脂垫上,在正置显微镜下进行拍照,使用Image J 2022软件测量每条线虫的面积和长度。实验重复3次,每个处理随机挑选30条线虫。
-
获得干净的线虫卵悬浮液后,确认卵浓度。暴露方法如1.2.4,选用无菌的96孔板,每孔约200枚卵,20 ℃恒温培养箱中暗处培养24 h,培养结束后在体式显微镜下计数卵和线虫的数量。
-
本实验以Davis团队的方法为标准,并在此基础上进行改善[35]。
-
1)挑出10条突变体菌株A1的L4雌雄同体线虫作为母本、CB4856雄虫30条作为父本,置于同1个NGM平板上,平板中央滴1滴OP50,20 ℃恒温培养。
2)培养24 h后,挑出10条雌雄同体的母本线虫(P0),单虫单板20 ℃恒温培养。
3)待母本(P0)产出F1代,观察F1代雄虫率,从雄虫率最高的P0板中挑出14条F1单虫单板培养。
4)待F1产出30~50条L1阶段的F2时,将14条F1分别进行裂解获得DNA裂解液,使用2对不同染色体的SNP位点引物进行PCR扩增,鉴定F1是否为杂合子线虫。
5)挑取鉴定为杂合子线虫的F1代中的子代,即F2代的L1阶段的线虫40条,使用1.2.1的测定方法进行表型验证,此过程需要筛选并获得杂合子F2。
6)72 h后将存活的F2代挑出250~300只,单虫单板培养。
7)待F2代产出30~50条L1阶段的F3代时,挑取F3代使用Waltheirone A在96孔板中进行表型验证,每孔30只,重复3次,48 h后,全部存活的则为突变纯合子,死亡率在100%的为野生型纯合子,死亡率在两者之间的为杂合子。
8)选择突变纯合子和杂合子单虫单板扩培,每个类别40个板。
-
使用SWL-蛋白酶K线虫DNA裂解液将挑选出的纯合子和杂合子F2代母虫单虫单管裂解,获得DNA,后将所有的杂合子DNA及纯合子DNA分别混成两管,将2管DNA分别制成PCR混液,酶切8 h后交替上样。
-
分析染色体定位结果,对染色体定位中SNP遗传标记位置进行区间定位,分别将突变纯合子单虫单管裂解成DNA,分别制成PCR混液,酶切8 h后上样。
-
首先进行线虫样品准备;从15个9 cm NGM培养板上收集临近饥饿期的秀丽隐杆线虫,用M9缓冲液冲洗并汇集至50 mL离心管。静置沉降后弃上清,将虫体转移至15 mL离心管,经2 500 r·min−1离心清洗2次去除培养基残留。接着用M9重悬后静置10 min,利用沉降差异去除细菌,吸弃上清后将沉淀转移至预称重的1.5 mL无酶EP管。最终用无酶水清洗虫体3次(12 000 r·min−1离心,4 ℃),彻底弃液后立即液氮速冻5 min,保存于−80 ℃并记录样品净重。后对秀丽隐杆线虫的突变株系(Mutant-A1)及野生型对照(N2)进行了全基因组测序;通过PacBio Sequel II平台获得高质量的HiFi长读段,并使用minimap2(v2.24, 参数:-ax map-hifi)软件将其高效比对至参考基因组(Caenorhabditis elegans, WBcel235)。随后,利用Clair3(v1.0.8)和GATK(v4.3.0)流程进行SNP与InDel的检测与联合基因分型,并使用VCFtools(v0.1.16, 参数:--min-meanDP 10 --maf 0.05 --max-missing 1)对结果进行严格过滤。最后,通过ANNOVAR对过滤后的变异进行功能注释,并利用定制Python脚本完成特定信息的提取。
-
根据TsingZol Total RNA Extraction Reagent TsingZol试剂盒说明书对秀丽隐杆线虫N2进行总RNA的提取。
-
使用HiScript III RT SuperMix for qPCR(+ gDNA wiper)试剂盒对总RNA进行反转录成cDNA,并对目的基因进行扩增。
-
对扩增后的目的基因条带进行切胶回收,纯化目的基因,后使用Xba Ⅰ及Sac Ⅱ对目的基因及质粒L4440进行双酶切获得粘性末端的目的基因及线性化质粒,后使用T4连接酶进行连接,最后将连接产物导入DH5α转化,将转化后的菌液涂抹在含有100 mg·L−1的氨苄青霉素(Amp)的LB固体培养基中过夜培养,最后挑取单菌落制成菌液,进行菌落PCR验证,验证后送去生工生物工程(上海)股份有限公司进行测序。
-
将测序匹配率达100%的菌液提取质粒,将目的基因-L4440及L4440导入HT115进行表达。构建完成的菌株在含有100 mg·L−1 Amp及6.25 mg·L−1 四环素(Tet)的LB培养基进行摇培,至OD600=0.4时加入IPTG至终浓度0.4 mmol·L−1,继续摇培4 h。对诱导后的大肠杆菌菌液进行总RNA提取,验证是否成功诱导dsRNA。
-
将诱导成功后的菌液饲喂于同步化至L1的野生型秀丽隐杆线虫N2,培育48 h后挑取F2的L1阶段的线虫进行Waltherione A敏感度检测,并洗下进行总RNA提取,进行RT-qPCR测定。
-
挑取40~50条F2的L1至96孔板中,以M9缓冲液为溶剂的10 mg·L−1的 Waltherione A处理组,同时设置空白对照组(仅含纯M9缓冲液)。所有处理均在20 ℃暗环境中暴露72 h后,统计死亡率。
-
分子对接验证从Pubchem(
https://pubchem.ncbi.nlm.nih.gov/ )数据库下载主要活性成分的分子结构,以sdf格式保存。从Alphafold数据库(https://alphafold.com/ )下载靶点蛋白的三维晶体结构,保存为PDB格式。使用PyMOL软件去除蛋白质的水分和配体保存为PDB格式,并使用Getbox Plugin插件获取对接口袋参数。将处理后的蛋白质和活性成分的文件分别导入AutoDock Tools 1.5.6转换为PDBQT格式。再使用AutoDock Vina 1.1.2执行分子对接。最后,用PyMOL 2.6.0软件和Discovery Studio 2019对分子对接结果进行2D可视化展示。 -
通过 IBM SPSS Statistics 19.0进行统计分析,对相关性指标在P<0.05 水平进行数据分析,采用t检验、单因素方差分析Tukey’s HSD进行显著性检验。用Origin 2022进行数据、图表处理,并制图分析。
-
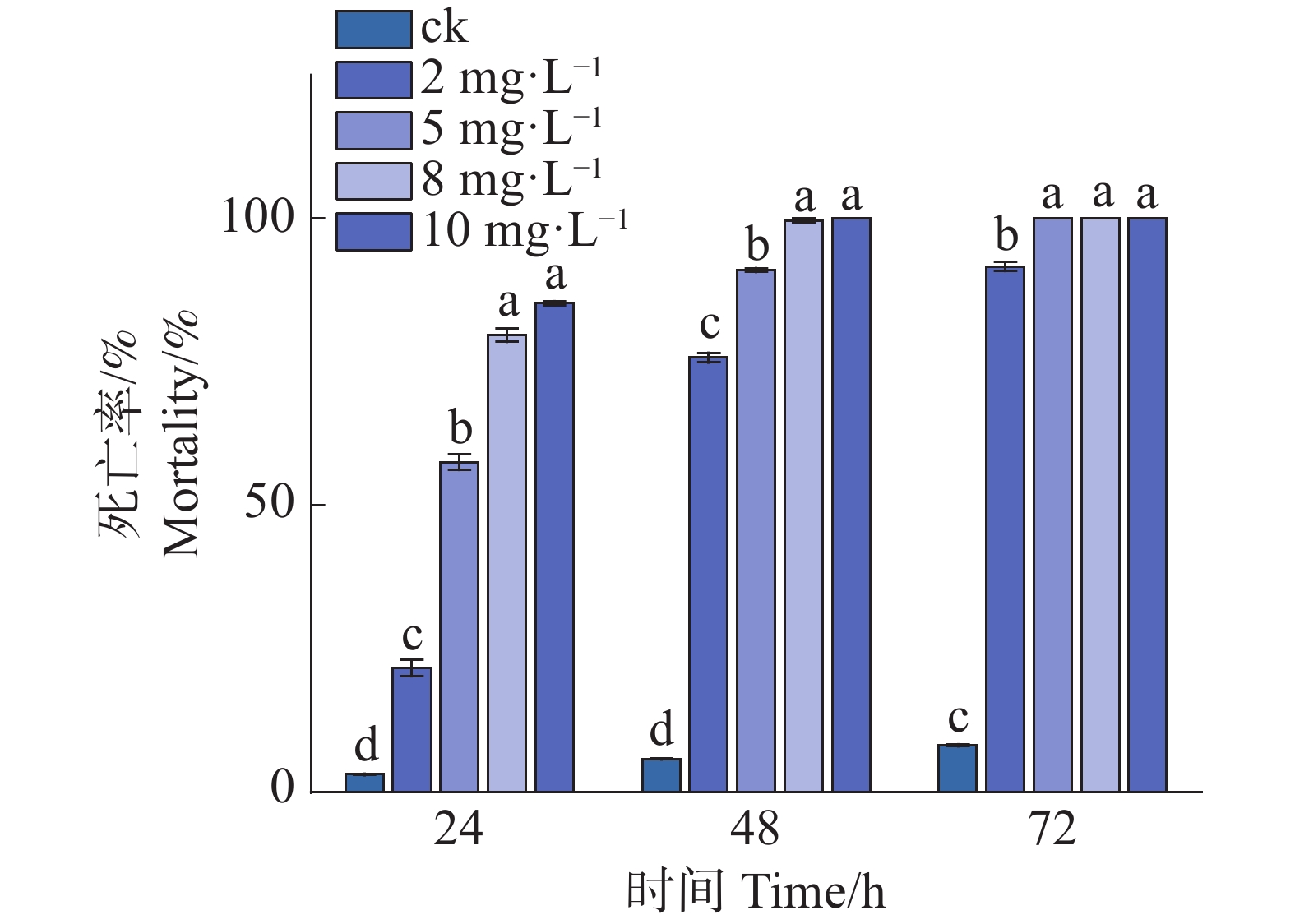
处理48 h后,8 mg·L−1组极少数线虫存活,而10 mg·L−1组均实现了100%的致死率(图1)。因此,在后续的突变体筛选及验证实验中,统一采用10 mg·L−1 Waltherione A作为处理质量浓度。
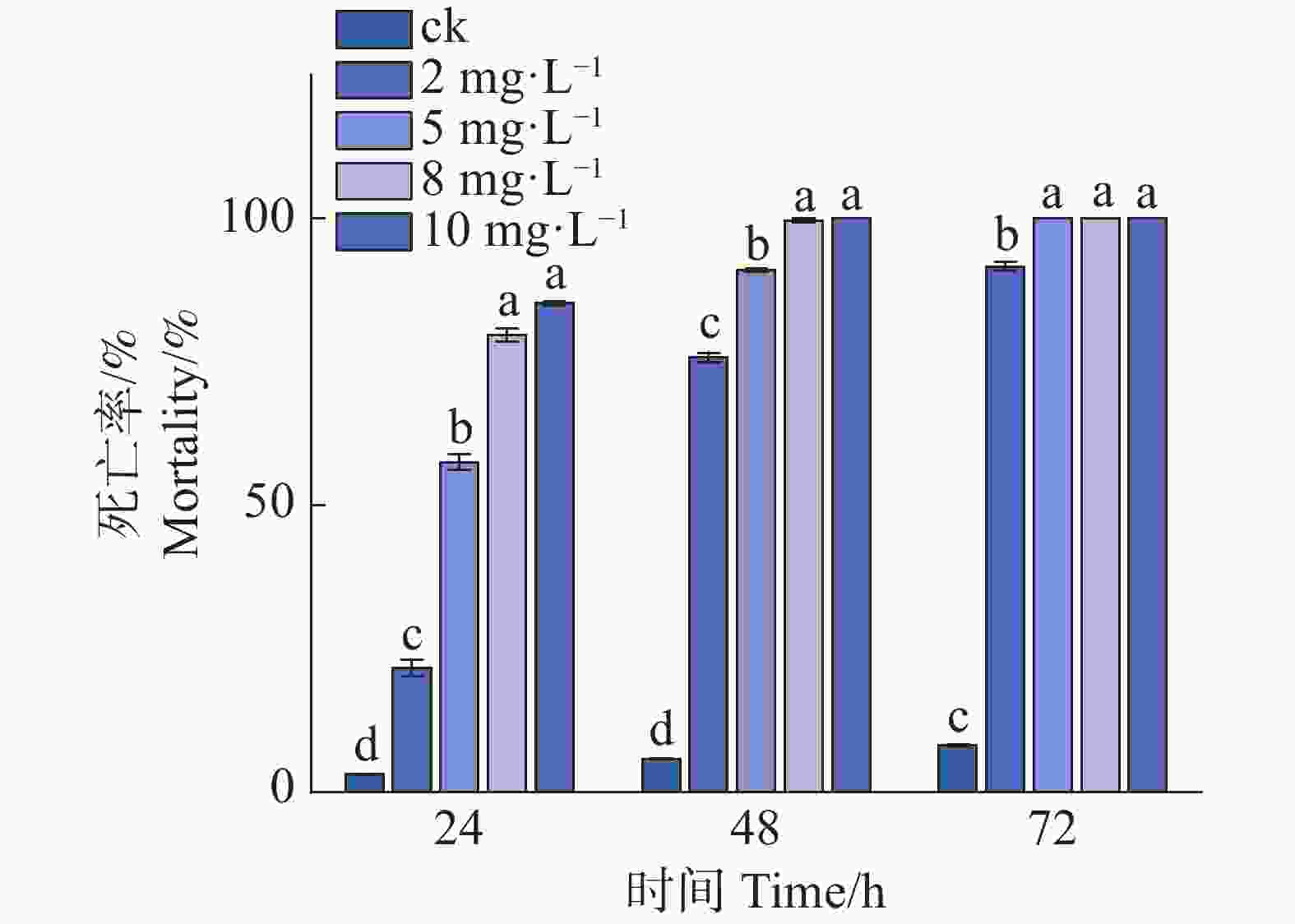
Figure 1. In vitro activity assay of Waltherione A against L1-stage larvae of the N2 strain
-
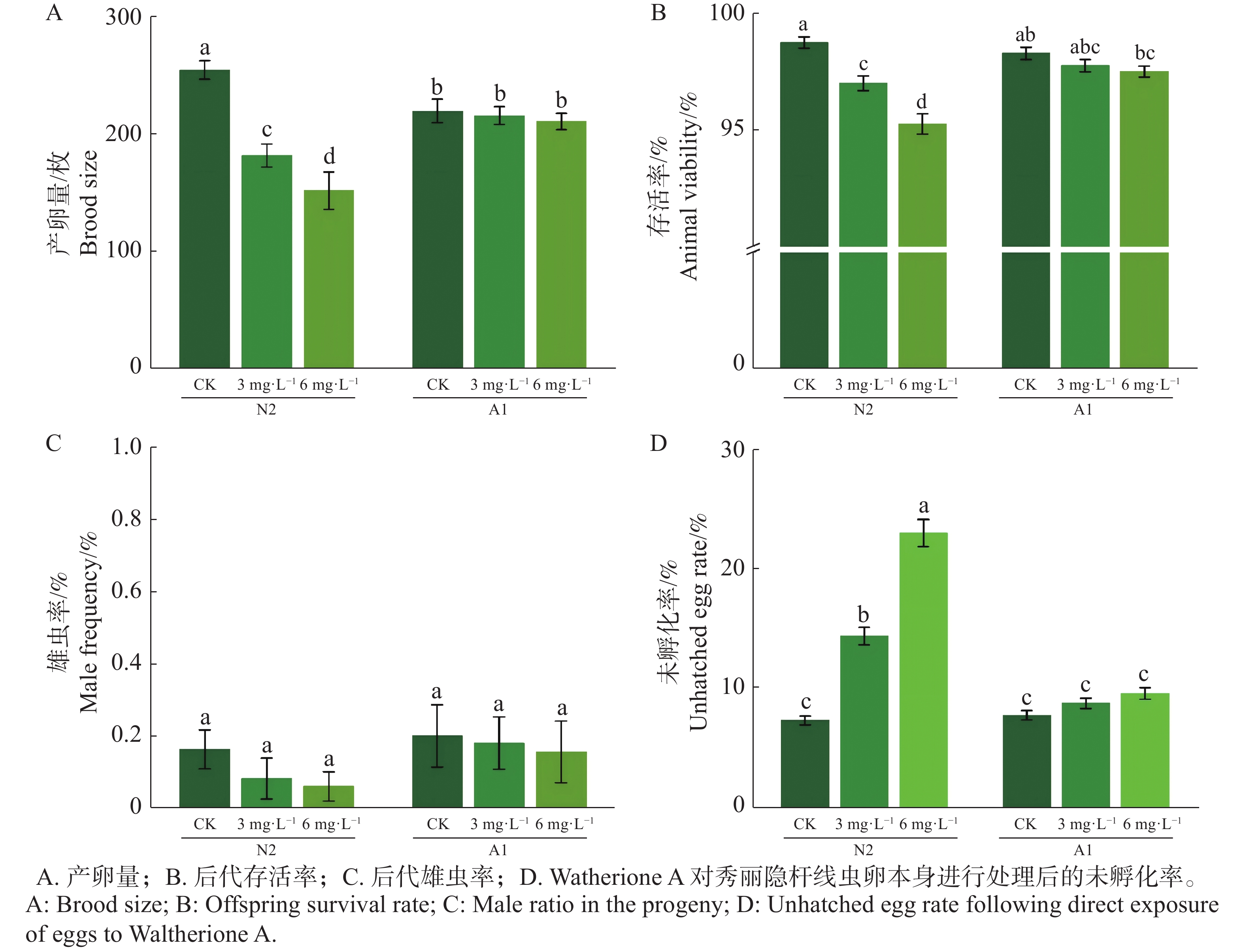
在对秀丽隐杆线虫L4进行0(CK)、3、6 mg·L−1 Waltherione A 处理 24 h(n = 15)条件下,野生型 N2 的繁殖输出呈显著剂量依赖性下降趋势;与对照组相比,野生型N2 的产卵量在 3 mg·L−1 和 6 mg·L−1 处理条件下分别下降28.63% 和 40.39%,后代存活率也随浓度升高也逐渐下降,呈现显著性差异,在后代雄虫率方面,各处理组雄虫比例均维持在正常水平,未有显著性差异(图2A-C)。而对野生型N2和突变体A1的卵悬浮液进行相同处理条件后,N2未孵化卵率由对照的7.26% 升高至14.30 % 和22.90 %,增加至对照的 2~3 倍(图2-D)。相比之下,抗性突变体 A1在相同处理条件下繁殖输出基本维持稳定,未孵化卵率仅小幅升高,显著低于 N2 对应处理组。
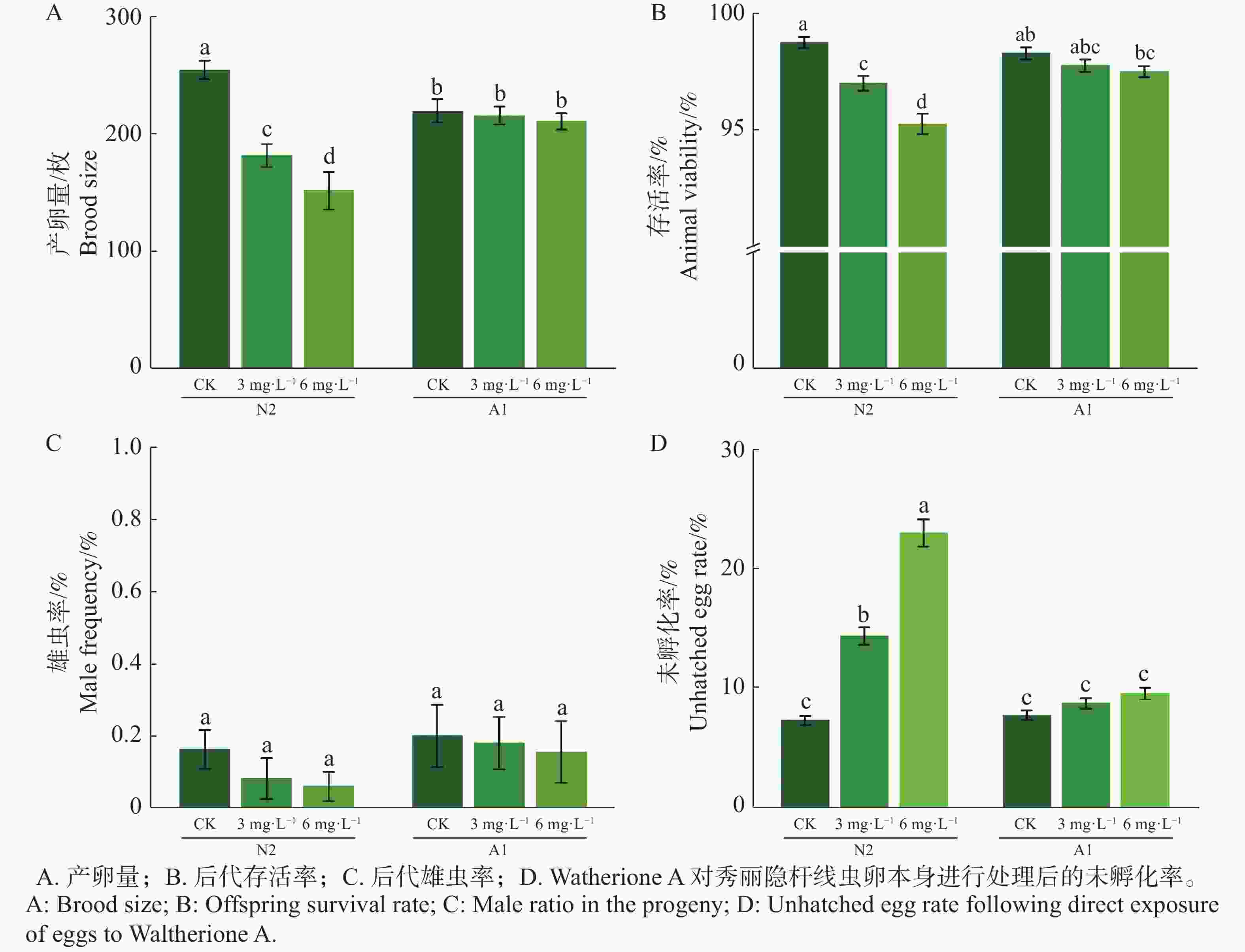
Figure 2. Effects of Waltherione A on the reproductive output of Caenorhabditis elegans
-
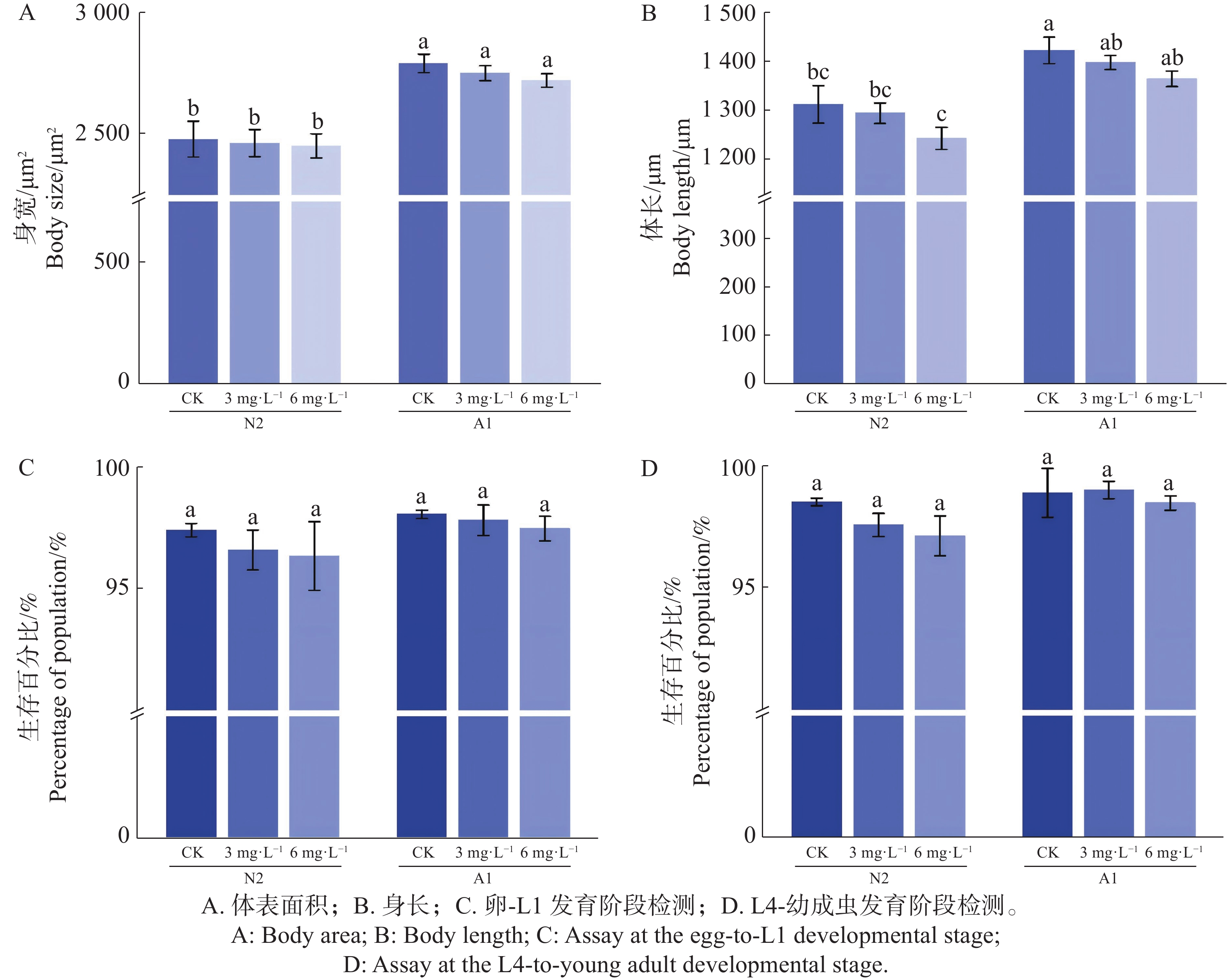
在对秀丽隐杆线虫L4进行0(CK)、3、6 mg·L−1 Waltherione A 处理 24 h(n=30)条件下,体表面积在野生型 N2 与 突变体A1 中均未随 Waltherione A 处理出现显著变化(图3-A);体长方面,N2 在 6 mg·L−1 条件下出现小幅缩短,而 A1 的体长变化不明显,但均未有显著差异(图3-B);在发育阶段监测方面,用Waltherione A处理后,N2、A1均能正常生长(图3-C—D)。表明 Waltherione A 在本实验条件下并未引起明显的整体生长发育抑制。
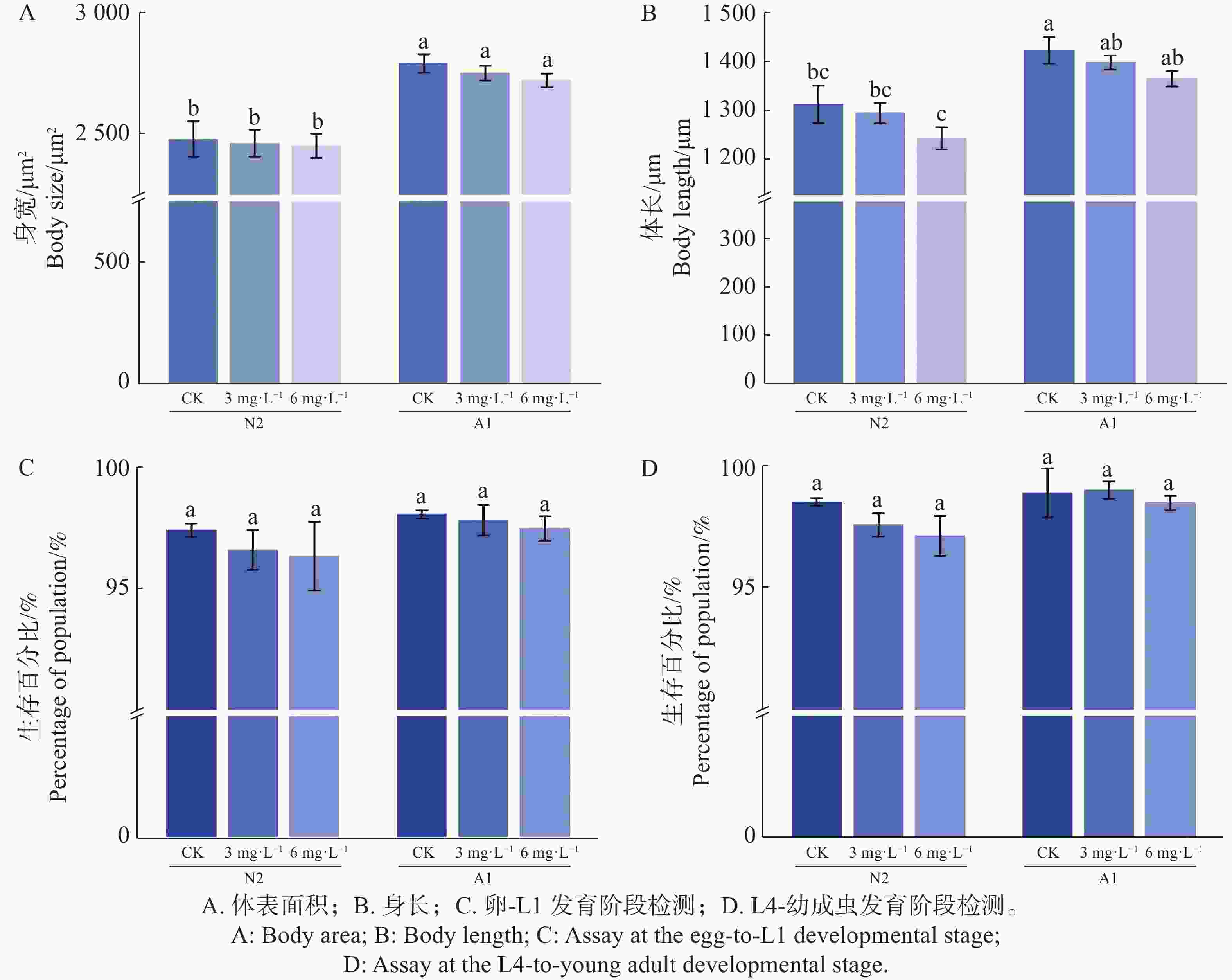
Figure 3. Effects of Waltherione A on the growth and development of Caenorhabditis elegans
-
夏威夷菌株CB4856雄性与突变体A1杂交,产生杂合F1代线虫,使用来自LG I (-19)和LG Ⅲ(-25)的引物对这些F1代线虫的DNA裂解液杂交基因型验证(图4-A)。基于突变表型(抗Waltherione A)鉴定来自杂合F1代线虫的纯合F2代线虫281条,一轮筛选得到31条待选突变体A1,使用10 µg·mL −1Waltherione A处理浓度进行复筛后得到36条纯合突变体A1。同时,还分离出了具有非纯合突变表型的线虫共40条。

Figure 4. SNP mapping results for mutant A1
-
经过系统性比对分析,染色体Ⅲ上的6个SNP标记(−12、−7、−1、4、12、21)显示出与非纯合突变泳道相比,纯合突变泳道中突变体A1 DNA的比例显著增加,这种连锁现象是明显的。观察到显著的条带富集差异(突变体A1型条带富集显著),这指示目标突变位点可能位于该染色体区域(图4-B)。
-
凝胶电泳图谱完整呈现了染色体Ⅲ上的6个SNP标记(−12,−7,−1,4,12,21)多态性带型,表明这些重组体在这些基因座是纯合的突变体A1,正如紧密连锁标记所预期的(图4-C)。从这些数据中,可以识别出含有目的基因的染色体,它们在Gene的左侧(重组型“−21”、“12”、“4”和“−1”)和Gene的右侧(重组型“−12”、“−7”)与夏威夷DNA重组。每个线虫含有2条含有基因的染色体,但在突变附近的区域,通常只有1条是重组的(图4)。正如预期的那样,定位数据将Gene放置在LGⅢ的−1和−7之间。指示目标突变位点可能定位于此区间范围(图4-D)。
-
通过连锁分析将A1的候选基因定位于LG Ⅲ遗传区间(−7,−1),对应物理位置F45H7至F56C9。基于全基因组测序,筛选出10个在突变体中均呈纯合突变的候选基因,包括4个编码区非同义突变和6个非编码区突变。在4个非同义突变中,F59A2(golg-4)、Y32H12A.4(szy-2)及F47D12.4(hmg-1.2)的突变位点位于已知功能结构域内,可能直接影响蛋白质功能;而B0280.7的突变结构域功能未知。其余6个基因Y42G9A.3、T27D1.3(npr-15)、F01F1.1(hpo-10)、T04C9.1、C13B9.4(pdfr-1)、C28A5.4(ceh-43)突变位于非编码区,可能通过调控表达影响表型,需进一步验证。
基因名称
Gene ID基因位点
Genelocus物理位置
Physical
Location原始序列
Raw seq变异序列
Change seq编码区突变
Mutation in coding regionF59A2.6
(golg-4)−6.96 3405638 GGCTGGATAATGAAGCCGGAAAGCTTAAACAATCAGCTCAGCAGTATGGAGAACAACTCGCCGCTCAAGTTGGACAATATC GGCTGGATAATGAAGCCGGAAAGCTTAAACAATCAGCTCAACAGTATGGAGAACAACTCGCCGCTCAAGTTGGACAATATC 是
(非同义
突变)T27D1.3
(npr-15)−4.82 3689753 GAATTAGAAATGGCTGAGCAAATAGGCAATTCGGTTGTTGGGGGTCGCGATGGAAGAAGAGCTGCTGGTTGCCACATAAAG GAATTAGAAATGGCTGAGCAAATAGGCAATTCGGTTGTTGAGGGTCGCGATGGAAGAAGAGCTGCTGGTTGCCACATAAAG 否 C28A5.4
(ceh-43)−3.17 4448125 GGTTCGGATTTTTTTTCGATCACGCTACTTTTAAATAATTGGTTTAGAAAATCCACACACAATTTTCCGGACACCCAATAC GGTTCGGATTTTTTTTCGATCACGCTACTTTTAAATAATTAGTTTAGAAAATCCACACACAATTTTCCGGACACCCAATAC 否 Y32H12A.4
(szy-2)−1.78 5371094 ATGCTTCCATAATCCTTGTCAGTTGGATGATAAGTGGCAAGAATATTCATTTCATCAAAATGTGCACGACCATCTTTGCCC ATGCTTCCATAATCCTTGTCAGTTGGATGATAAGTGGCAAAAATATTCATTTCATCAAAATGTGCACGACCATCTTTGCCC 是
(非同义
突变)F01F1.1
(hpo-10 )−1.44 5879902 TTCCAGTGCCAATGGAGACAGGAGCAAATTCCGAAGCTTGGAAAAAAGCGAATGAAGCATTAATGAAGGTTTGTAAATATT TTCCAGTGCCAATGGAGACAGGAGCAAATTCCGAAGCTTGAAAAAAAGCGAATGAAGCATTAATGAAGGTTTGTAAATATT 否 T04C9.1 −1.43 6002197 GTCTTACTTTTCACTTTACACCCCCGGCAATCGATCCCATGGGAATCTGGTAGGGGACTCAATGGACAAAGTTGAATGATC GTCTTACTTTTCACTTTACACCCCCGGCAATCGATCCCATAGGAATCTGGTAGGGGACTCAATGGACAAAGTTGAATGATC 否 Y42G9A.3 −1.41 6147905 CGGAGTTCCATCAGTCGACTACGATACTTCGTCTGAATCAGAAGAGGAGATTGAGCCAATCATGAGAAGAGTCTTGCTTGG CGGAGTTCCATCAGTCGACTACGATACTTCGTCTGAATCAAAAGAGGAGATTGAGCCAATCATGAGAAGAGTCTTGCTTGG 否 F47D12.4
(hmg-1.2)−1.41 6281641 CTTCTTTTACTCACAAGACAAACGTCCTGAAATACAGGCAGGTCATCCAGATTGGAAAGTCGGACAAGTTGCACAGGAGCT CTTCTTTTACTCACAAGACAAACGTCCTGAAATACAGGCAAGTCATCCAGATTGGAAAGTCGGACAAGTTGCACAGGAGCT 是
(非同义
突变)C13B9.4
(pdfr-1 )−1.03 6639646 CTCAAAAAATTTTATTGCAAAACGGTCACGTTTGTAGAATCAGGTAAAAATTGATCAAATGTTTGTATCGAAATCACTATT CTCAAAAAATTTTATTGCAAAACGGTCACGTTTGTAGAATTAGGTAAAAATTGATCAAATGTTTGTATCGAAATCACTATT 否 B0280.7 −0.79 7111323 AAAATGACAGTTCCATTACTGAGACTTCTTGTCTCTCCTGGATGCATTATTTCTCCTTGTGTATCAATACAAGCAATCGCT AAAATGACAGTTCCATTACTGAGACTTCTTGTCTCTCCTGAATGCATTATTTCTCCTTGTGTATCAATACAAGCAATCGCT 是
(非同义
突变)Table 1. Information about candidate genes of non-synonymous mutations in A1 mutants
-
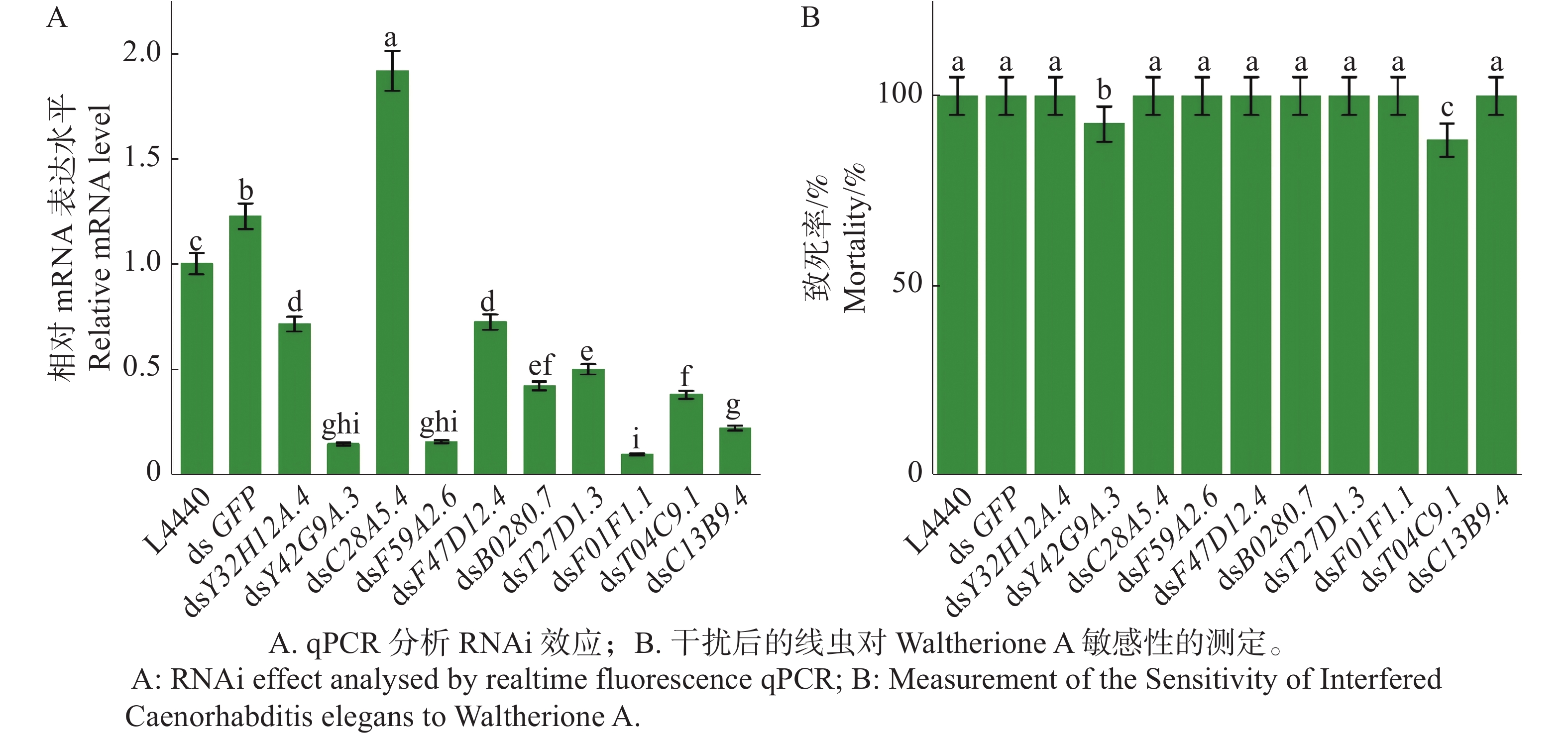
Y42G9A.3、F59A2.(golg-4)、B0280.7、T27D1.3(npr-15)、F01F1.1(hpo-10 )、T04C9.1、C13B9.4(pdfr-1 )被成功低敲,而Y32H12A.4(szy-2)和F47D12.4(hmg-1.2)低敲效果不明显,而C28A5.4(ceh-43)反而过表达(图5 -A)。接着对被成功低敲的目标基因的野生型秀丽隐杆线虫N2进行Waltherione A敏感性测定;F59A2.(golg-4)、B0280.7、T27D1.3(npr-15)、F01F1.1(hpo-10 )、C13B9.4(pdfr-1 )、Y32H12A.4(szy-2)、F47D12.4(hmg-1.2)被低敲后均对Waltherione A敏感,而Y42G9A.3和T04C9.1被低敲后秀丽隐杆线虫对Waltherione A产生了弱抗性(图5-B)。
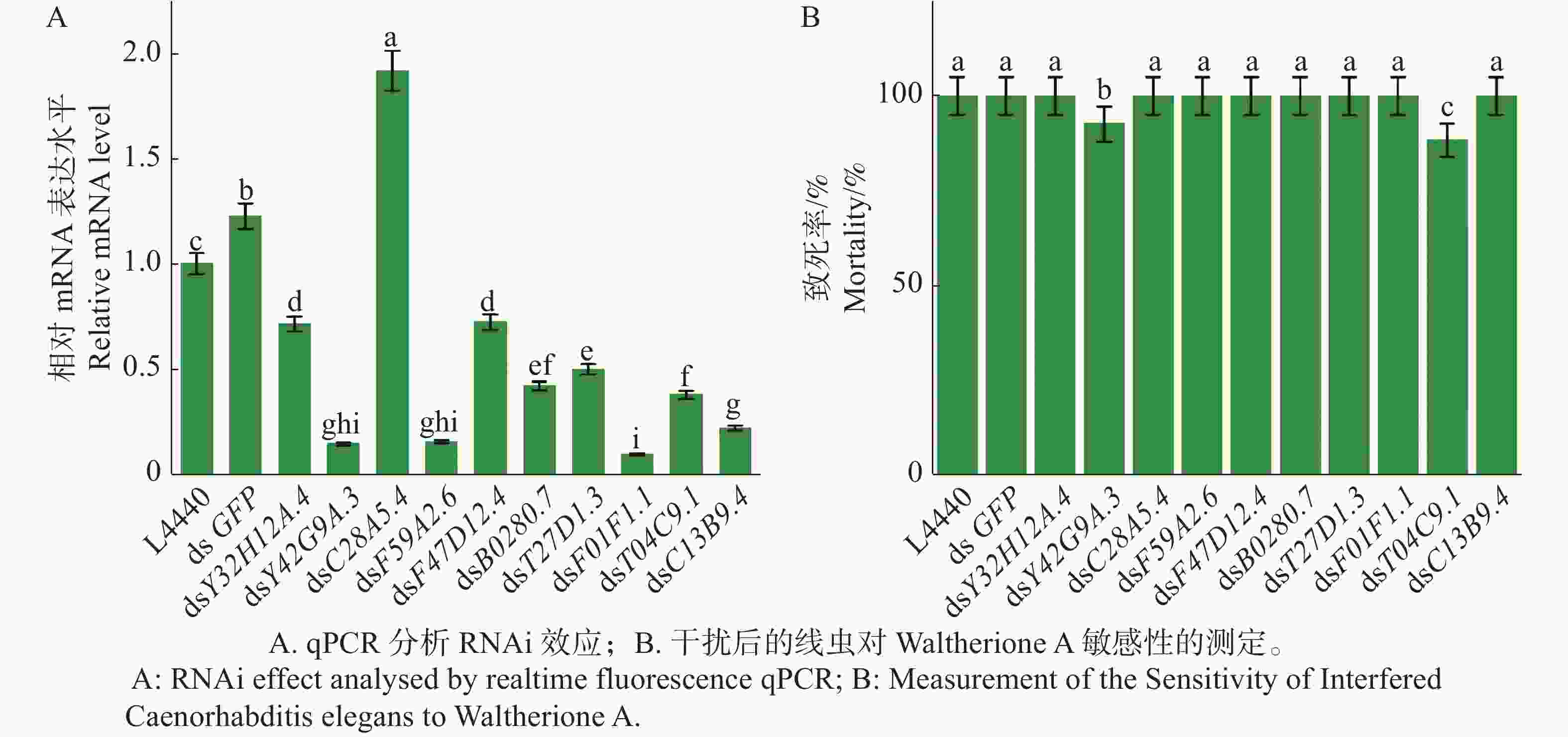
Figure 5. SNP Mapping results for mutant A1
-
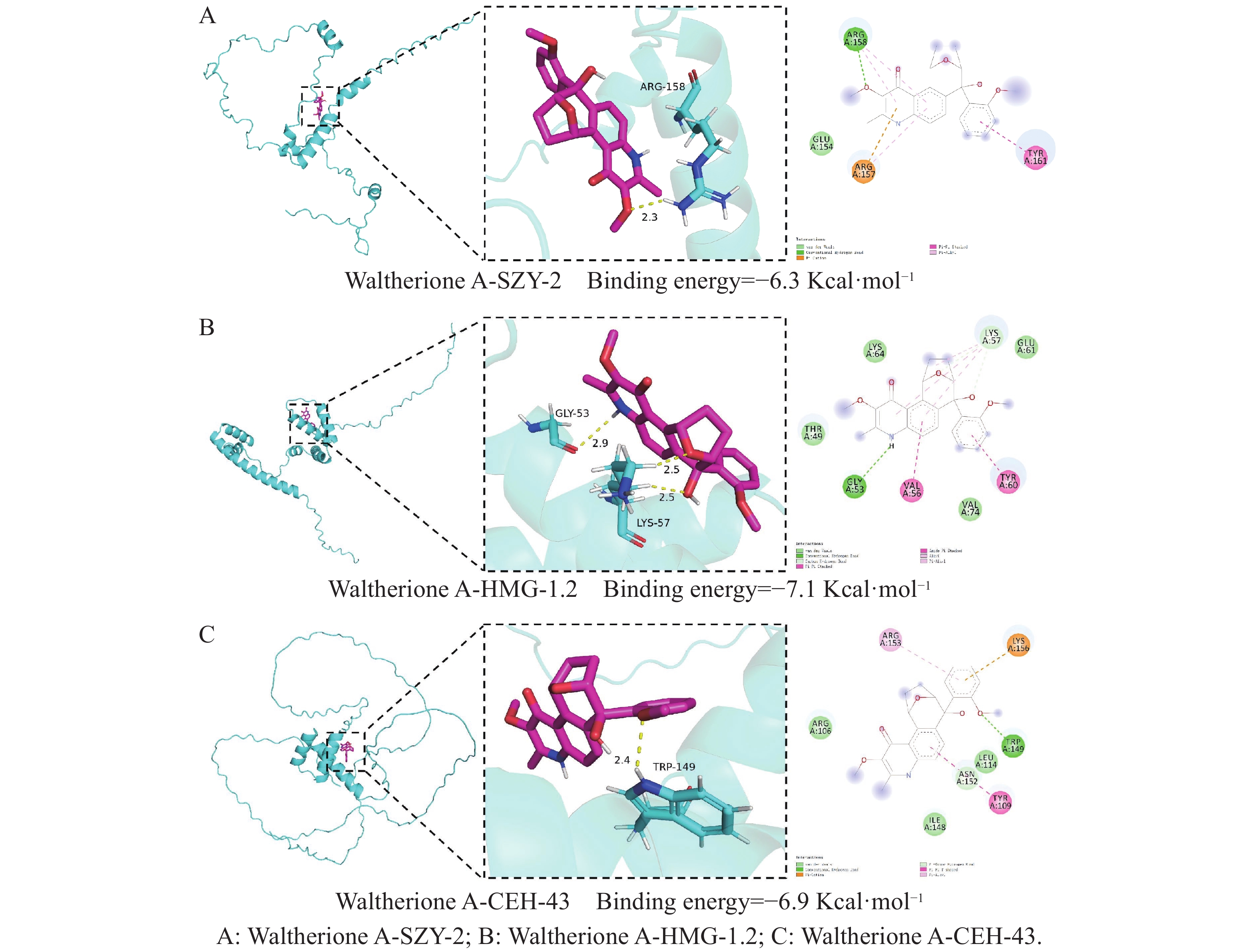
SZY-2(由Y32H12A.4表达)与Waltherione A的结合能为−6.3 kcal·mol−1(图6-A);HMG-1.2(由F47D12.4表达)与Waltherione A的结合能为−7.1 kcal·mol−1(图6- B);CEH-43(由C28A5.4表达)与Waltherione A的结合能为−6.9 kcal·mol−1(图6- C)。由此得出Waltherione A能与Y32H12A.4(szy-2)、F47D12.4(hmg-1.2)和C28A5.4(ceh-43)所表征的蛋白在理论上均存在自发且稳定的结合潜力。
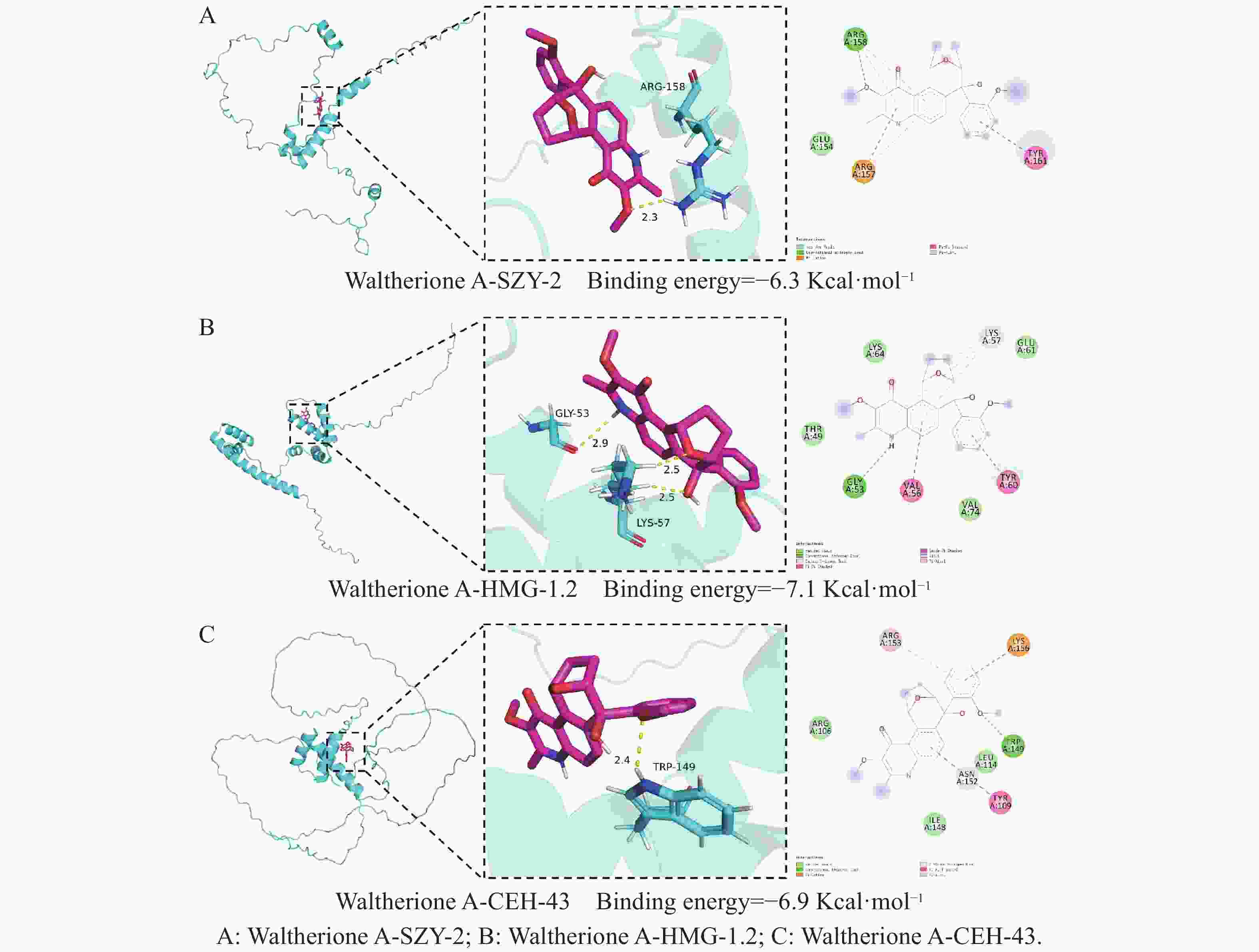
Figure 6. Moleculardocking results
-
随着农业系统面临愈加严峻的根结线虫危害,亟需开发具有全新作用机制的天然来源杀线剂。目前,大多数商业化杀线剂仍集中于有限的经典靶点,如线粒体复合体 I(fluopyram)、谷氨酸门控氯离子通道(阿维菌素)、乙酰辅酶 A 羧化酶(spirotetramat),这不仅提高了抗性出现的速度,也削弱了田间的持续控制效果[36-37]。与此同时,根结线虫复杂的寄生适应能力及土壤中长期存活特性,使传统化学防控在可持续农业背景下愈发力不从心[38]。因此,系统解析天然产物的杀线机制,尤其是靶向新型生理节点的化合物,对农业防控具有重要意义。
本研究通过系统的活性测定、基因定位与功能验证,初步揭示了天然化合物Waltherione A在线虫中的作用机制。Waltherione A在10 mg·L−1质量浓度下48 h对野生型线虫N2的L1具有完全致死效果,并呈现显著的生殖毒性,表现为产卵量、后代存活率及卵孵化率的剂量依赖性下降,而对体长、体表面积等基础生长发育参数影响有限。表明,Waltherione A的毒性机制可能特异性靶向于生殖和早期胚胎发育相关的精密调控通路,而非广泛的生长发育抑制。
利用SNP mapping技术,将决定抗性的突变位点定位于连锁群Ⅲ(LG III)上的−7与−1之间的遗传区间。结合全基因组重测序,在该区间内鉴定出10个潜在靶标基因。
随后的RNAi实验提供了最直接的功能性证据:由于Y42G9A.3和T04C9.1被低敲后产生的抗性极其微弱(死亡率未达到50%),所以可能是源于这2个基因本身所获得的抗药性,并非Waltherione A的关键靶点。因此,排除了以上被低敲成功后的所有基因。由此推测,未被成功低敲的Y32H12A.4(szy-2)、F47D12.4(hmg-1.2)和C28A5.4(ceh-43)可能是Waltherione A杀线虫作用机制中关键通路的关键作用基因。在Wormbase等数据库中查找到Y32H12A.4基因所表征的蛋白是SZY-2,此蛋白作为蛋白磷酸酶的关键调控蛋白,以磷酸化依赖的方式精确控制其活性,从而参与细胞信号传导、代谢、细胞周期等多种核心生命过程;而HMG-1.2(由F47D12.4表达)是线虫中高迁移率族蛋白家族的成员,属于染色质相关的核蛋白,其核心功能是作为转录调控因子:它能直接结合DNA(即具有“转录顺式调节区结合活性”),通过参与Wnt信号通路、促进RNA聚合酶II的转录等过程,从染色质结构和基因表达层面调控多种生理过程(如外阴发育等);在Aspöck[39]的研究结果表明,CEH-43(由C28A5.4表达)是线虫中1个关键的发育调控转录因子,其主要功能是调控胚胎早期前端结构的正常形成与细胞间的粘附,该蛋白在特定前皮下细胞(如Hyp3、Hyp4)及CAN等神经元中表达,其功能缺失会导致严重的胚胎发育缺陷:包括前感觉凹陷处细胞泄漏、咽部无法连接口腔并缩回体内、尾部变形以及表皮脆弱易破裂,这些表型表明,CEH-43对于胚胎前端皮下细胞的正确发育、细胞间粘附力的维持以及后续口咽结构的正常连接至关重要,是保障胚胎形态发生和结构完整性的必需因子。综合实验数据与生物信息学分析推测,Waltherione A的杀线虫作用或许并非通过单一靶点实现,而可能涉及1个由SZY-2、HMG-1.2和CEH-43协同构成的调控网络。该化合物可能通过直接干扰SZY-2(蛋白磷酸酶关键调控蛋白)所介导的细胞信号与周期进程,同时影响HMG-1.2(染色质关联转录因子)所调控的基因表达程序,从而在线虫体内产生多重胁迫。这种协同干扰或许会进一步异常调动CEH-43(发育关键转录因子)所主导的胚胎发育与细胞粘附通路,最终导致线虫稳态崩溃与死亡。此多靶点协同干扰的假设,为从信号传导、转录调控到发育执行层面系统解析其分子机制,提供了新的、可验证的研究方向。
分子对接模拟从理论上支持了相互作用的可能性,计算显示Waltherione A能与这3个蛋白模型自发结合,其中与HMG-1.2的结合能(−7.1 kcal·mol−1)最为突出,预示其结合可能更稳定。遗传学表型与计算化学预测的相互印证,加强了这3个基因作为直接或关键下游效应分子的可信度。
基于此,本研究提出1个创新性的整合作用模型:Waltherione A可能并非作用于单一孤立的靶点,而是通过协同干扰1个多层次网络来发挥毒性—即干扰HMG-1.2介导的全局转录调控,同时影CEH-43控制的特定发育程序,并可能通过SZY-2破坏下游的信号传导与细胞周期进程。该网络的失调,可能最终导致线虫发育与稳态崩溃并死亡。当然,这一模型仍需后续实验严格验证,包括使用等温滴定量热或表面等离子共振技术证实化合物与蛋白的直接物理互作,以及通过条件性基因敲除等更精细的遗传操作确认其功能必要性。
-
本研究遵循“从表型到基因”的正向遗传学思路,逐步揭示并勾勒出Waltherione A潜在的作用路径。通过验证该化合物的毒性特征集中于生殖系统;通过SNP mapping与全基因组测序,将潜在抗性基因定位至LG III区间,并得到候选基因。最后,通过RNAi功能筛选与分子对接模拟,分析得到Y32H12A.4(szy-2)、F47D12.4(hmg-1.2)和 C28A5.4(ceh-43)为介导Waltherione A毒性的3个关键基因。这一发现创新性地提出了Waltherione A可能通过协同干扰1个涉及信号传导、基因表达与发育程序的多层次网络来发挥杀线活性的新假说,为从分子层面完整揭示其作用机制提供了至关重要的实验线索与明确方向,也为创制基于新作用模式的绿色杀线剂奠定了理论基础。
Phenotype and garget gene screening in a Waltherione A-resistant Caenorhabditis elegans mutant
DOI: 10.15886/j.cnki.rdswxb.20260019
- Received Date: 2026-01-21
- Accepted Date: 2026-02-11
- Rev Recd Date: 2026-02-11
- Available Online: 2026-02-28
-
Key words:
- Waltherione A /
- Caenorhabditis elegans /
- resistant mutant /
- RNAi.
Abstract: Waltherione A is a plant-derived nematicidal compound with significant activity against root-knot nematodes (LC50 0.09–3.54 mg·L−1). Caenorhabditis elegans was used as a model organism to elucidate the action mechanism of Waltherinone A. Based on a pre-determined screening concentration of 10 mg·L−1, phenotypic differences between wild-type N2 and the Waltherione A-resistant mutant A1 were compared at treatment concentrations of 3 and 6 mg·L−1, enabling a basic phenotypic characterization of the mutant. Systematic screening via SNP mapping and whole-genome resequencing, combined with RNAi validation, identified three potential target genes: Y32H12A.4 (szy-2), F47D12.4 (hmg-1.2), and C28A5.4 (ceh-43). Molecular docking simulations indicated that Waltherione A can form stable, high-affinity complexes with the corresponding proteins (binding energies of −6.3, −7.1, and −6.9 kcal·mol−1, respectively). The results suggest that Waltherione A may exert its nematicidal effect by simultaneously targeting the transcription regulator HMG-1.2, the developmental programming factor CEH-43, and the phosphatase signaling protein SZY-2, thereby disrupting a coordinated response pathway governed by transcriptional and developmental regulation with phosphatase signaling as a key execution node. All these findings systematically reveal the multi-target network of Waltherione A, providing an important theoretical basis for its further development and mechanistic application as a novel green nematicide.
| Citation: | Fu Yingjie, Bai Yiongxia, Wang Shuai, Sun Ranfeng. Phenotype and garget gene screening in a Waltherione A-resistant Caenorhabditis elegans mutant[J]. Journal of Tropical Biology. doi: 10.15886/j.cnki.rdswxb.20260019

|
 DownLoad:
DownLoad: