-
铜绿假单胞菌(Pseudomonas aeruginosa,P. aeruginosa)是经常引发医院获得性感染的机会性致病菌,在6种常见的人类威胁性病原细菌(ESKAPE)中被评为最高级[1]。P. aeruginosa易感染患有肺囊纤维化、慢性肺阻塞、烧伤的患者和使用植入医疗器械或长期服用抗菌药物的患者;在免疫缺陷人群中感染P. aeruginosa的发病情况尤为严重,如每年都会有几百万的菌血症感染甚至死亡案例发生[2-3]。铜绿假单胞菌的环境适应性极强,在中国的海南、广东等热带与亚热带地区高温、高湿环境中易定植和传播,例如其污染桶装饮用水的事件已严重威胁食品安全和公共卫生[4-7]。P. aeruginosa丝状噬菌体(P. aeruginosa filamentous bacteriophages,Pf)是一类侵染铜绿假单胞菌的温和噬菌体;Pf能维持生物被膜完整性,从而显著地增强细菌的致病性,进而增强细菌的抗生素耐药性并延长细菌感染的持久性[8-10]。因此,研究Pf噬菌体对于防治P. aeruginosa感染具有潜在的临床意义。
丝状噬菌体Pf属丝杆病毒,呈纤细的长丝状,由蛋白外壳包裹一条单链DNA组成[11]。Pf分布在60%以上临床或环境菌株中[10],Pf能在细菌群落的P. aeruginosa菌体间流通并整合到基因组上[12]。Pf4是感染P. aeruginosa实验室标准菌株PAO1的温和噬菌体。Pf4既能以成熟的噬菌体颗粒形式存在,也能整合细菌基因组的甘氨酸tRNA编码基因(pa0729.1)的下游形成原噬菌体,还能从基因组切离形成细菌体胞内的小型双链环状DNA并以此为模板进行复制和转录表达。丝状噬菌体Pf4编码的切离酶XisF4是促使Pf4从宿主细菌基因组切离并复制的核心调控蛋白[13]。XisF4除介导原噬菌体切离之外,还能作为转录调控因子,结合于多个噬菌体基因启动子区进行调节转录表达[13]。
Tang M等[14]2024年发现,XisF4的调控作用不仅局限于原噬菌体Pf4,还能影响宿主细菌基因组上多个基因的表达,但切离酶XisF4的蛋白理化性质及其调控作用机制还需要更深入地解析,因此,本研究选取临床环境中最常见的丝状噬菌体Pf4编码的切离酶XisF4进行研究,提供了XisF4在调控铜绿假单胞菌临床感染的作用机制方面的科学新认识。
-
使用Expasy在线工具ProtParam(
https://web.expasy.org/protparam/ )分析XisF4蛋白的氨基酸组成、相对分子质量、理论等电点(pI)、不稳定系数、亲水性总平均值(GRAVY)等基本理化性质。采用SOPMA(https://npsa-prabi.ibcp.fr/ )预测其二级结构组成。通过AlphaFold3(https://alphafoldserver.com/ )进行从头建模,获取XisF4蛋白的三级结构模型,并获得预测TM分数(Predicted TM-score);同时也进行XisF4与DNA复合物的从头建模,分析预测的关键结合位点,并获得预测局部距离差测试分数(pLDDT)。使用TMHMM 2.0(https://services.healthtech.dtu.dk/services/TMHMM/ )预测跨膜结构域,SignalP 5.0(https://services.healthtech.dtu.dk/services/SignalP/ )预测信号肽。通过MEME Suite(https://meme-suite.org/meme/ )对已知启动子序列进行de novo motif预测,识别XisF4特异性结合序列。 -
XisF4编码序列经高保真DNA聚合酶Phanta UniFi(南京诺唯赞)PCR扩增后(引物见表1),克隆至经EcoRI(美国赛默飞世尔)和HindIII(美国赛默飞世尔)双酶切处理的pHERD20T表达载体(来源于本实验室前期已发表工作[14]),构建重组质粒pHERD20T::xisF4。转化至E. coli DH5α感受态细胞(上海碧云天)进行扩增,经DNA聚合酶Taq酶反应体系(南京诺唯赞)进行菌落PCR及Sanger测序(上海生工生物)验证序列正确性。将验证正确的重组质粒经电转化至经300 mmol·L−1蔗糖溶液(上海麦克林)多次洗涤并重悬制备的P. aeruginosa PAO1 ΔPf4感受态细胞(来源于本实验室前期已发表工作[14]),以终浓度20 mmol·L−1 L-阿拉伯糖(上海麦克林)为诱导剂,诱导XisF4蛋白表达5 h。
引物名称 引物序列(5′—3′) Primer Sequnce(5′—3′) pHERD1 CTCTCTACTGTTTCTCCATACCCGT pHERD2 GCTGCAAGGCGATTAAGTTGGGTAA XisF4-pHERD1 gtaaaacgacggccagtgccaagcttCTTGCAATCCATAATGCGTAATTAG XisF4-pHERD2 AGTGAAAAGTTCTTCTCCTTTACGCATGCTCATTCCGTCACTATTCGTGG pet-sumo-xisF4F GGCTCACAGAGAACAGATTGGTGGTATGGAACTGGAAGAGCTTAG pet-sumo-xisF4R GGCTTTGTTAGCAGCCGGATCTCATCAGGCACTCCATTCCTGTTC pet-sumo-verify01 GCTGATCAGACCCCTGAAGA pet-sumo-verify02 CCCCAAGGGGTTATGCTAGT 16SRNA-F CCCAACATCTCACGACACGA 16SRNA-R ACGCGAAGAACCTTACCTGG xisF4-qF CCGCAACAGGATGTGGAG xisF4-qR GCACTCCATTCCTGTTCCAA pqsB1 CTGATCGAGAACCGCGACAT pqsB2 TCGGTGAGGTCGAAGACATG hcpC-F ACCGTCCTGGAAGACGCGAT hcpC-R AGTGCACGTCTTCCAGGTGG phzB2 TGCGCCGCCATGAACTTTTC phzB2 CAGCTTGTCCTTGCCACGAA Table 1. Primer sequence information
-
收集上述诱导XisF4表达后的菌体,超声破碎提取总蛋白;转化pHERD20T空载体的菌体作为对照组,每组3个重复。蛋白粗提液经4℃,5 000 r·min−1离心20 min并通过0.45μm孔径的无菌滤头(美国默克密理博)。使用BCA法(上海生工生物)测定蛋白浓度,经SDS-PAGE质检合格后,进行胰酶酶解。肽段采用液相色谱-串联质谱(LC-MS/MS)技术进行数据非依赖采集(DIA)。所有样本均使用spike-in的iRT肽段进行保留时间校准,以确保跨批次色谱保留时间的一致性。原始数据通过Spectronaut™软件(Biognosys)进行数据库搜索与定量分析。在所有定量蛋白质中,技术重复间的中位变异系数(coefficient of variation,CV)小于15%,这表明定量结果具有高度可重复性。在蛋白质和肽段水平均应用严格的数据依赖性和谱图库搜索策略,并使用SpectronautTM默认的Q值进行质量控制。差异表达蛋白(DEPs)的筛选标准为差异倍数(Fold Change)≥1.5,且经多重检验后的校正P值(adjusted P-value)<0.05。使用GO、KEGG数据库进行功能注释与通路富集分析,STRING数据库构建蛋白互作网络(PPI)。蛋白组组学数据的质谱分析数据已归档至蛋白组资源平台iProx(
https://www.iprox.cn/ ),该项目编号为IPX0014531000。 -
诱导XisF4蛋白表达5 h后,依照RNA提取试剂盒(常州百代生物)流程指示提取细菌RNA,并使用逆转录试剂盒(南京诺唯赞)将RNA逆转录为cDNA。利用Primer5设计定量引物,内参基因为16S rRNA(引物见表1)。通过实时荧光定量PCR(RT-qPCR)获取数据,验证XisF4在过表达状态下潜在靶基因的转录水平变化。两组间基因表达量的显著性差异由t检验统计计算。
-
XisF4编码序列经高保真DNA聚合酶Phanta UniFi(南京诺唯赞)PCR扩增后(引物见表1),克隆至经EcoRI(美国赛默飞世尔)和XhoI(美国赛默飞世尔)双酶切处理的pET28a表达载体,经过热激转化进入大肠杆菌BL21(DE3)感受态细胞(北京全式金)。复苏后将菌液涂布于含50 mg·L−1卡那霉素的LB固体培养基平板,37℃过夜培养后挑取单菌落进行PCR验证,将阳性克隆子送至生工生物工程(上海)股份有限公司进行测序。
将含有pET-28a-xisF4重组载体的大肠杆菌BL21(DE3)菌株接种至含有50 mg·L−1卡那霉素的LB液体培养基中,在37℃摇床培养至OD600在0.4~0.6之间,加入IPTG至终浓度为100 μmol·L−1,在150 r·min−1、15℃条件下,过夜诱导表达12 h。然后将菌液冷冻离心(4℃,4 500 r·min−1)20 min,用10 mL的Tris-HCl(50 mmol·L−1 Tris,350 mmol·L−1 NaCl)缓冲液重悬菌体。超声破碎仪设置为超声时间4 s,间隙时间4 s,总时间20 min,冰浴条件下(温度4℃左右),功率40%。冰上超声破碎重悬菌体后将破碎的菌体冷冻离心(4℃,12 000 r·min−1)10~15 min,分离得到沉淀和上清液。提前取出镍柱,用20 mL Tris-HCl缓冲液清洗2~3遍后关闭镍柱阀门,将破碎上清液经0.45 μm孔径的无菌滤头(美国默克密理博)过滤后注入镍柱中并封口,置于冰上150 r·min−1孵育1 h。然后收集孵育上清液,用20 mmol·L−1咪唑洗脱杂蛋白,300 mmol·L−1咪唑收集目的蛋白XisF4。最后进行SDS聚丙烯酰胺凝胶电泳(SDS-PAGE)分析。
-
潜在调控靶点基因xisF4、pa0727、phz2、hcpC启动子序列进行高保真PCR(南京诺唯赞)扩增,并进行PCR产物纯化(北京天根)。使用5×EMSA/Gel-Shift结合缓冲液(上海碧云天),将浓度梯度递增的XisF4蛋白与各启动子DNA片段在室温下孵育20 min,随后进行聚丙烯酰胺凝胶电泳(PAGE),经GelRed核酸染料(上海生工生物)染色后成像分析。
-
从NCBI数据库下载XisF4、XisF5及衣壳蛋白CoaA编码基因的同源序列,使用MEGA 11.0软件进行多序列比对(ClustalW)。采用最大似然法(Maximum Likelihood)构建系统发育树,Bootstrap值设置为
1000 次重复以评估节点支持率。通过比较不同蛋白的进化树拓扑结构,分析进化关系与进化规律。 -
XisF4的蛋白分子式为C362H574N100O106S2,包含71个氨基酸,相对分子质量为8 087.2,总原子数为
1144 ,理论等电点为5.32。XisF4蛋白的氨基酸组成中丙氨酸(Ala)和精氨酸(Arg)含量最高(均为8.5%);带负电荷的氨基酸残基(Asp+Glu)为9,带正电荷的氨基酸残基(Arg+Lys)为8(表2);不稳定系数为46.33,属于不稳定蛋白(标准≦40的为稳定蛋白);280 nm处的吸光系数(A280)是24 980 L·mol−1·cm−1;脂肪系数平均为100.28,总平均亲水性为−0.235。氨基酸
Amino acid数目
Number占比/%
Proportion/%氨基酸
Amino acid数目
Number占比/%
Proportion/%Ala(A) 6 8.5 Lys(K) 2 2.8 Arg(R) 6 8.5 Met(M) 2 2.8 Asn(N) 2 2.8 Phe(F) 0 0.0 Asp(D) 1 1.4 Pro(P) 3 4.2 Cys(C) 0 0.0 Ser(S) 7 9.9 Gln(Q) 3 4.2 Thr(T) 2 2.8 Glu(E) 8 11.3 Trp(W) 4 5.6 Gly(G) 5 7.0 Tyr(Y) 2 2.8 His(H) 0 0.0 Val(V) 5 7.0 Ile(I) 3 4.2 Pyl(O) 0 0.0 Leu(L) 10 14.1 Sec(U) 0 0.0 Table 2. Amino acid composition of the protein
-
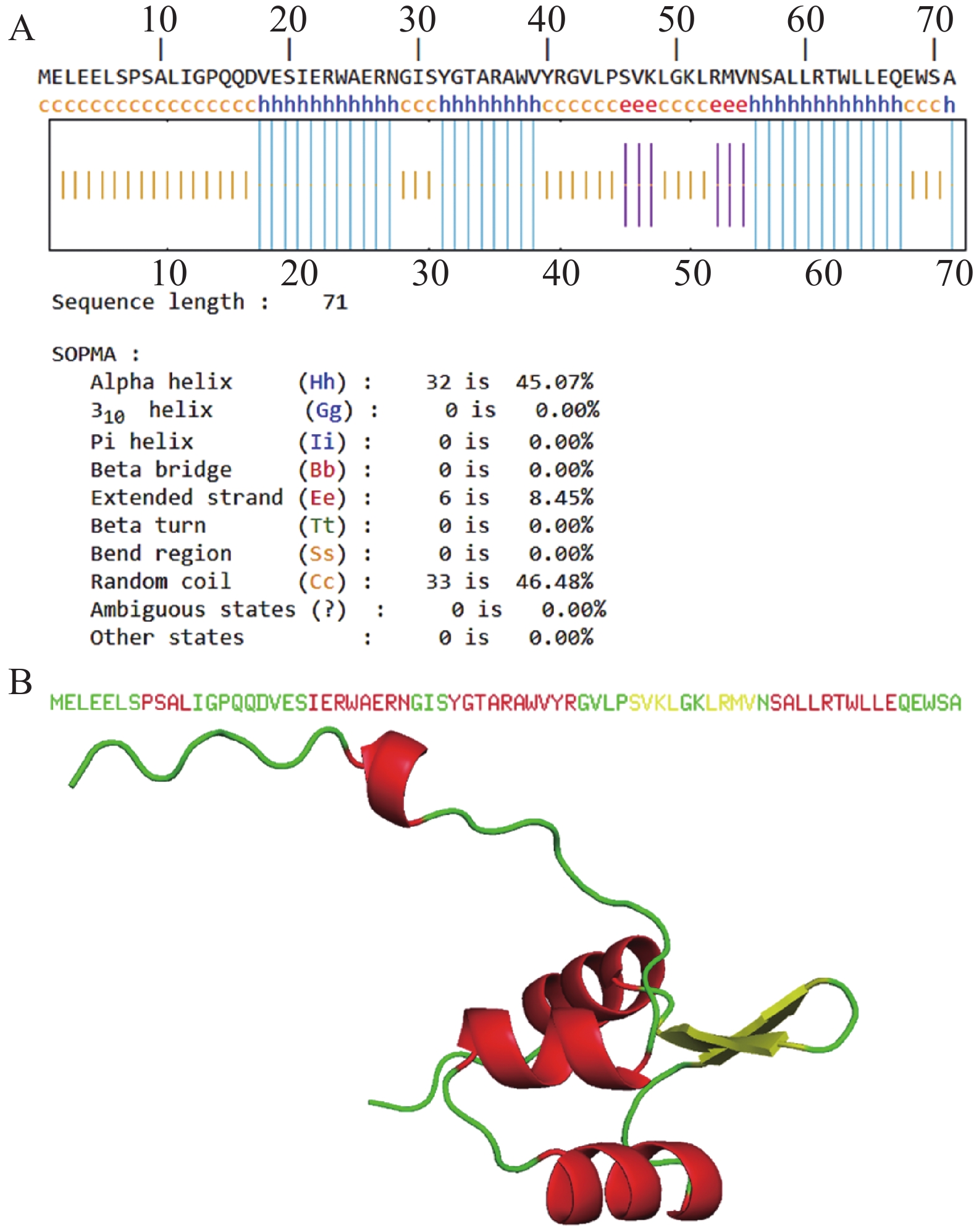
XisF4蛋白中有32个氨基酸参与形成α螺旋,占总氨基酸比例为45.07%;6个氨基酸参与延伸链的形成,占总氨基酸比例为8.45%;33个氨基酸参与无规卷曲结构的形成,占总氨基酸比例为46.48%;0个氨基酸参与β折叠的形成(图1-A)。AlphaFold3对XisF4蛋白三级结构进行同源建模。pTM值可用于初步评估预测模型的质量,其值越接近1表明建模质量越好。模型预测XisF4的pTM值为0.69,表明该预测模型质量可靠。图1-B为XisF4蛋白三级结构的从头建模结果,其以α螺旋和无规则折叠为主,与二级结构预测结果相吻合。
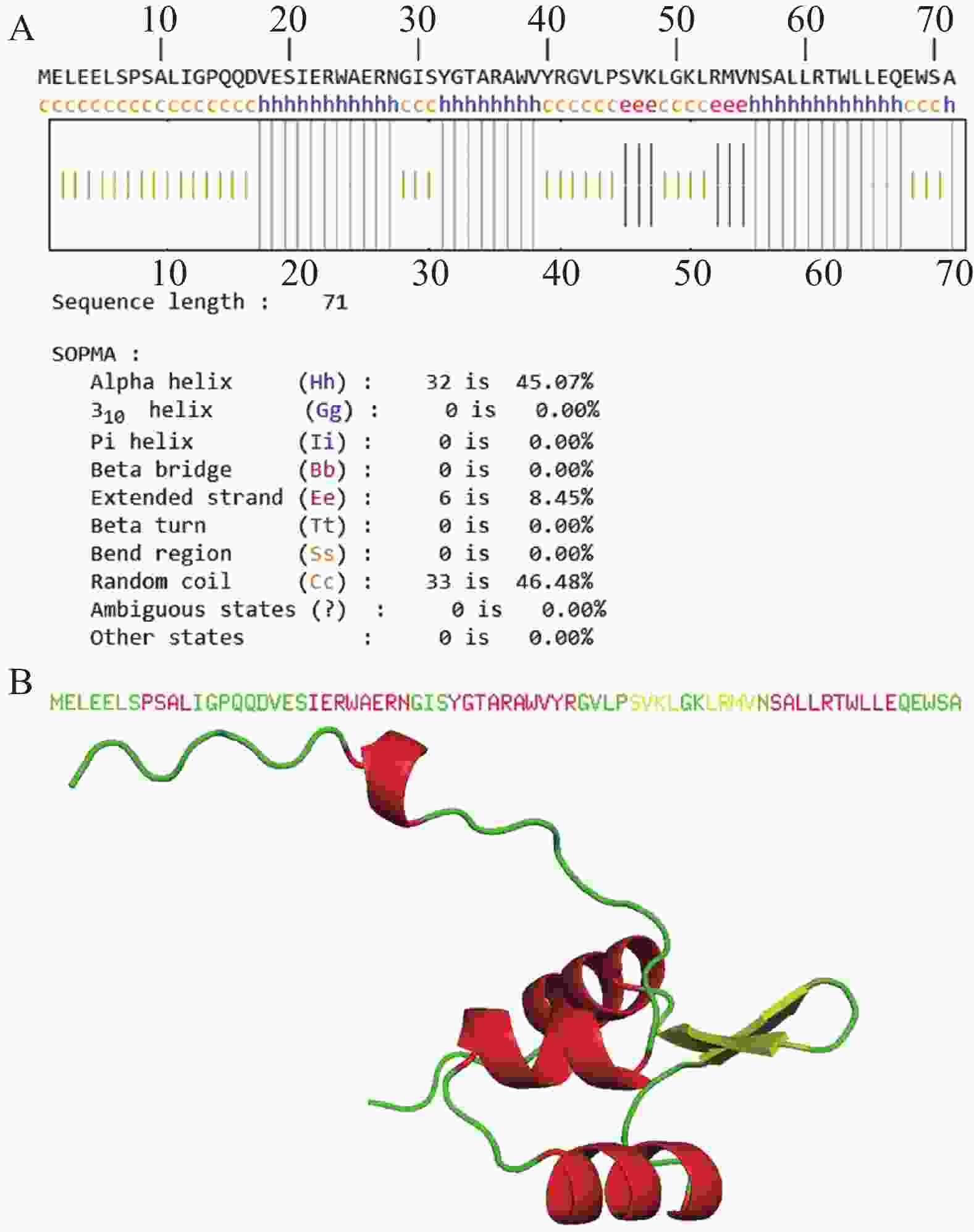
Figure 1. Prediction of XisF4 protein secondary (A)and tertiary (B)structures
-
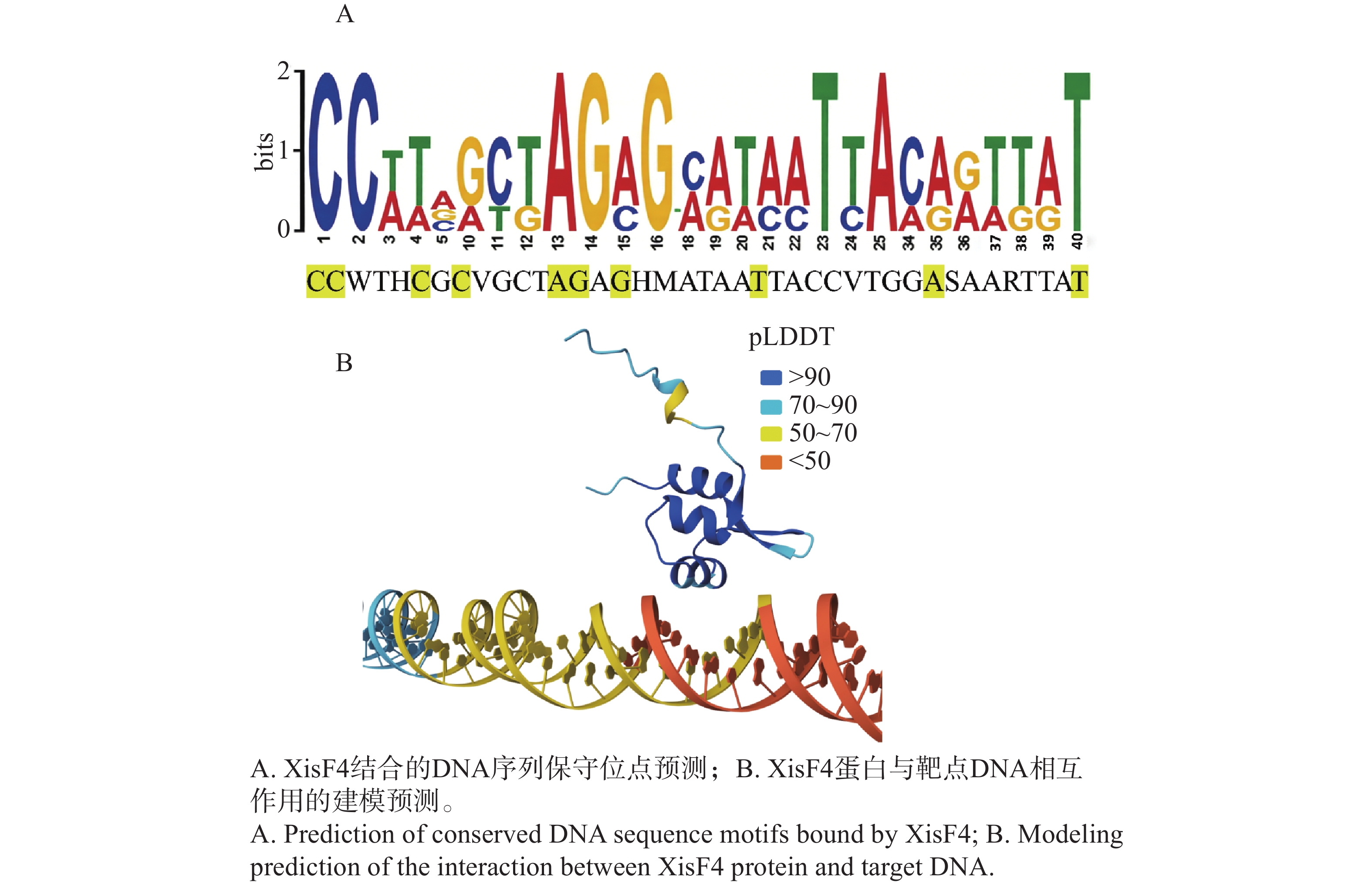
TMHMM预测XisF4没有跨膜结构域,且信号肽的概率极低(0.475%),提示XisF4是一个胞内蛋白质,这与XisF4作为DNA结合的切离酶及转录调控因子的特性相一致。XisF4的第20位至第64位氨基酸序列组成了典型的属于MerR型超家族的螺旋—转角—螺旋DNA结合结构域(图2-A)。为了探索噬菌体切离酶XisF4的关键氨基酸位点在进化上的保守性,XisF4及同源蛋白的氨基酸序列进行排列对比。结果表明XisF4的第24、29、36、38、42、45、48、50、51、54、55、56位氨基酸残基高度保守,且全部位于该螺旋—转角—螺旋DNA结合结构域内,提示这些位点可能是识别和结合DNA的关键氨基酸位点(图2-B)。此外,XisF4与P. aeruginosa种内典型MerR家族蛋白的氨基酸序列进行了进一步对比。结果表明XisF4的第29、36、42、48、54位氨基酸残基在MerR家族蛋白其他蛋白成员中也高度保守(图2-C),更加佐证了这几个氨基酸位点在蛋白识别与结合DNA过程中的关键作用。Tang等[14]及其他研究团队的既往电泳迁移率变动分析EMSA实验先后发现,XisF4能与xisF4、pf4r、intF4和pilA四个基因的启动子区域结合[13-14]。因此,这些启动子序列被进一步对比分析,由此预测到XisF4能结合的DNA保守序列为CCWTHCGCVGCTAGAGHMATAATTACCVTGGASAARTTAT(图3-A)。基于以上结果,XisF4蛋白与DNA保守序列的相互作用进行了AlphaFold3预测建模。pLDDT值用于评估模型对每个氨基酸位置的预测置信度,其分数范围从0到100,数值越高表示预测的结构越可靠,通常用于评估蛋白质结构中每个位置的预测准确度。建模结果再次提示XisF4的20位至64位氨基酸位点区域为DNA结合结构域(图3-B),其中第32~41位氨基酸残基组成的α螺旋是最可能靠近或插入DNA深沟的区域。

Figure 2. Analysis of functional domain and conserved amino acid residues of XisF4 protein
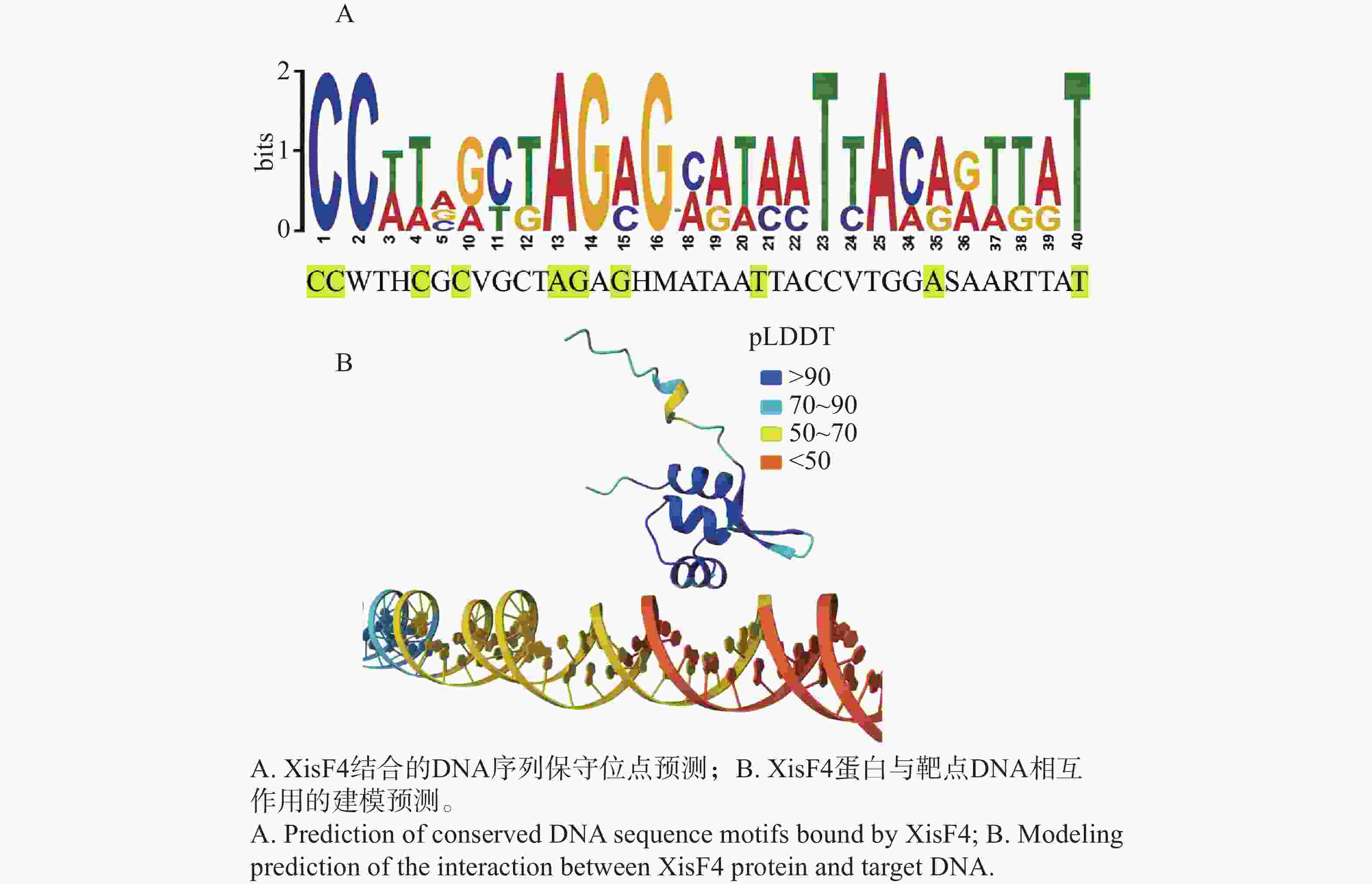
Figure 3. Predicted DNA conservative sequence bound by XisF4
-
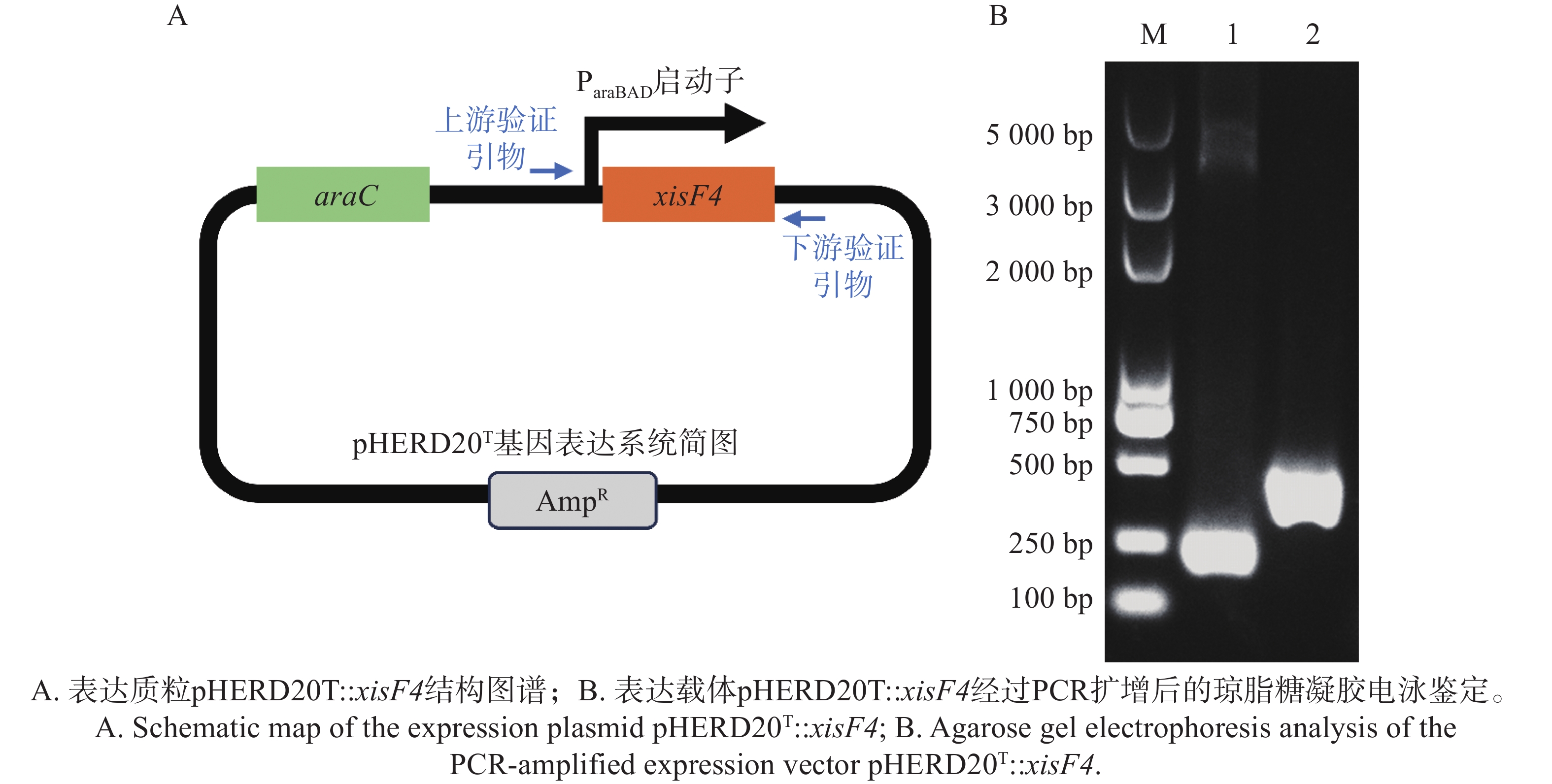
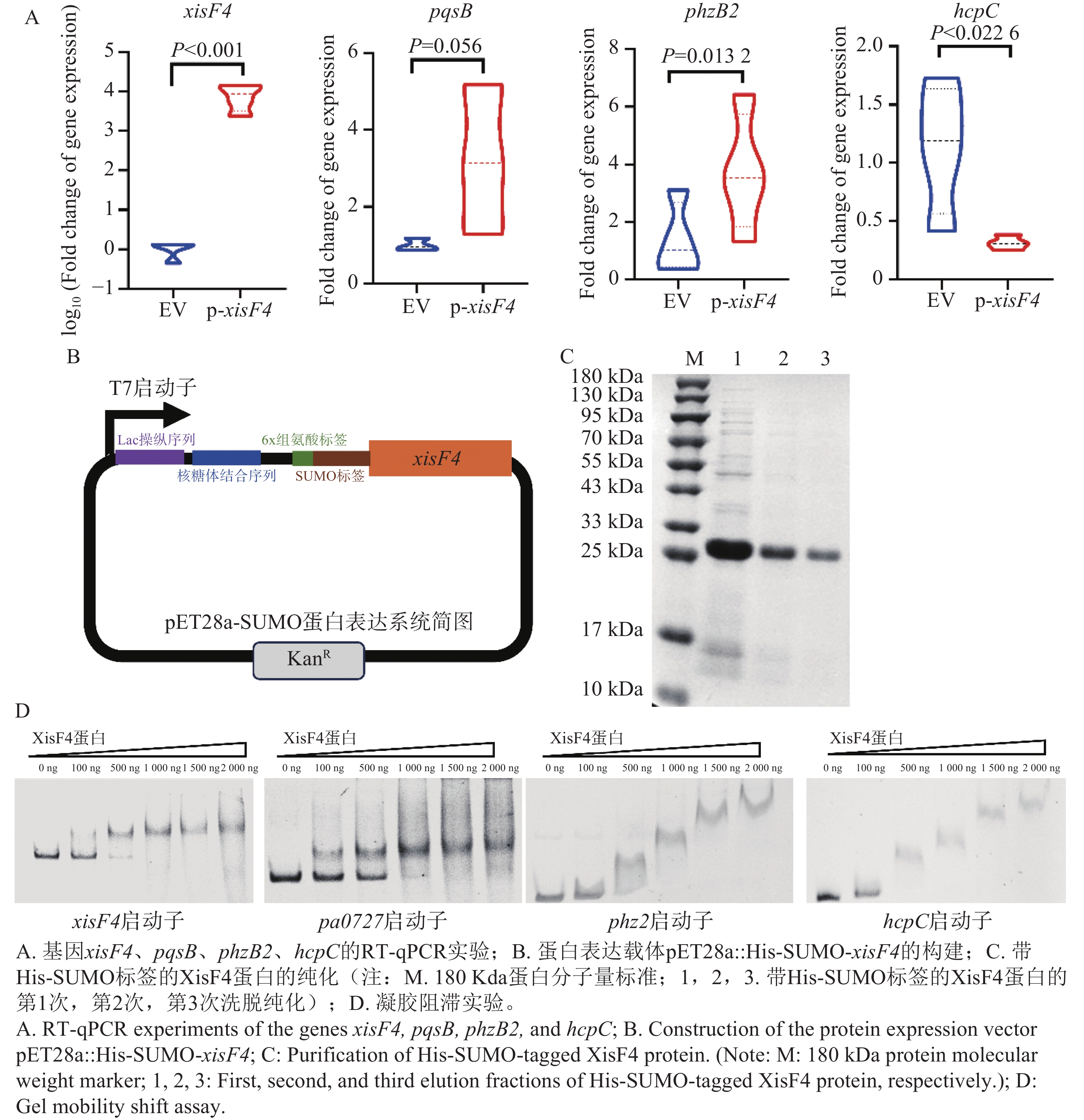
XisF4蛋白编码区域经高保真PCR扩增并连接至经限制性内切酶EcoRI和HindIII消化后呈线型的表达质粒pHERD20T上,得到表达质粒pHERD20T::xisF4(图4-A),并最终经高保真PCR反应和Sanger测序确认目标表达载体质粒序列无误(图4-B)。
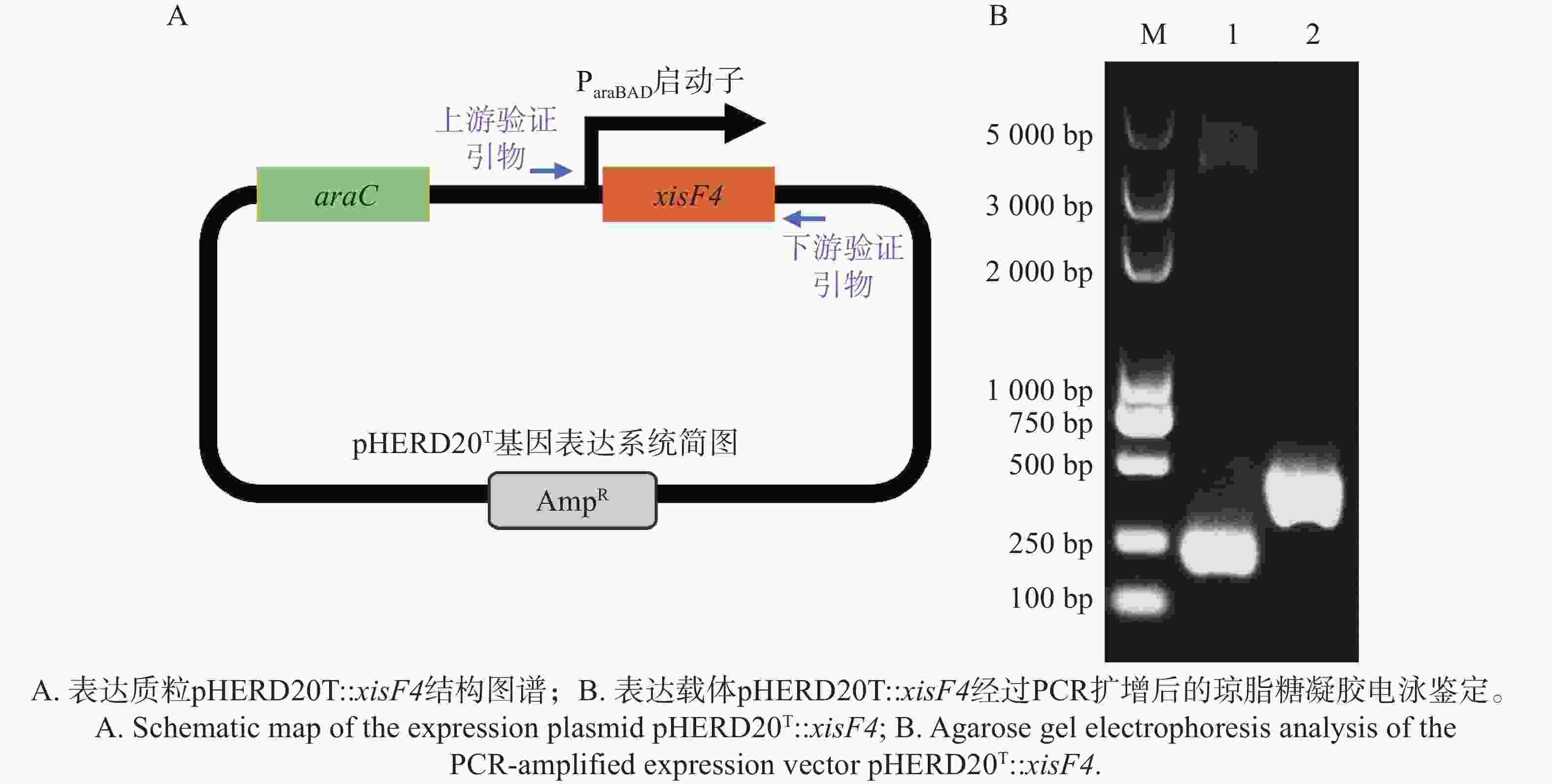
Figure 4. Construction and identification of the expression plasmid pHERD20T::xisF4
-
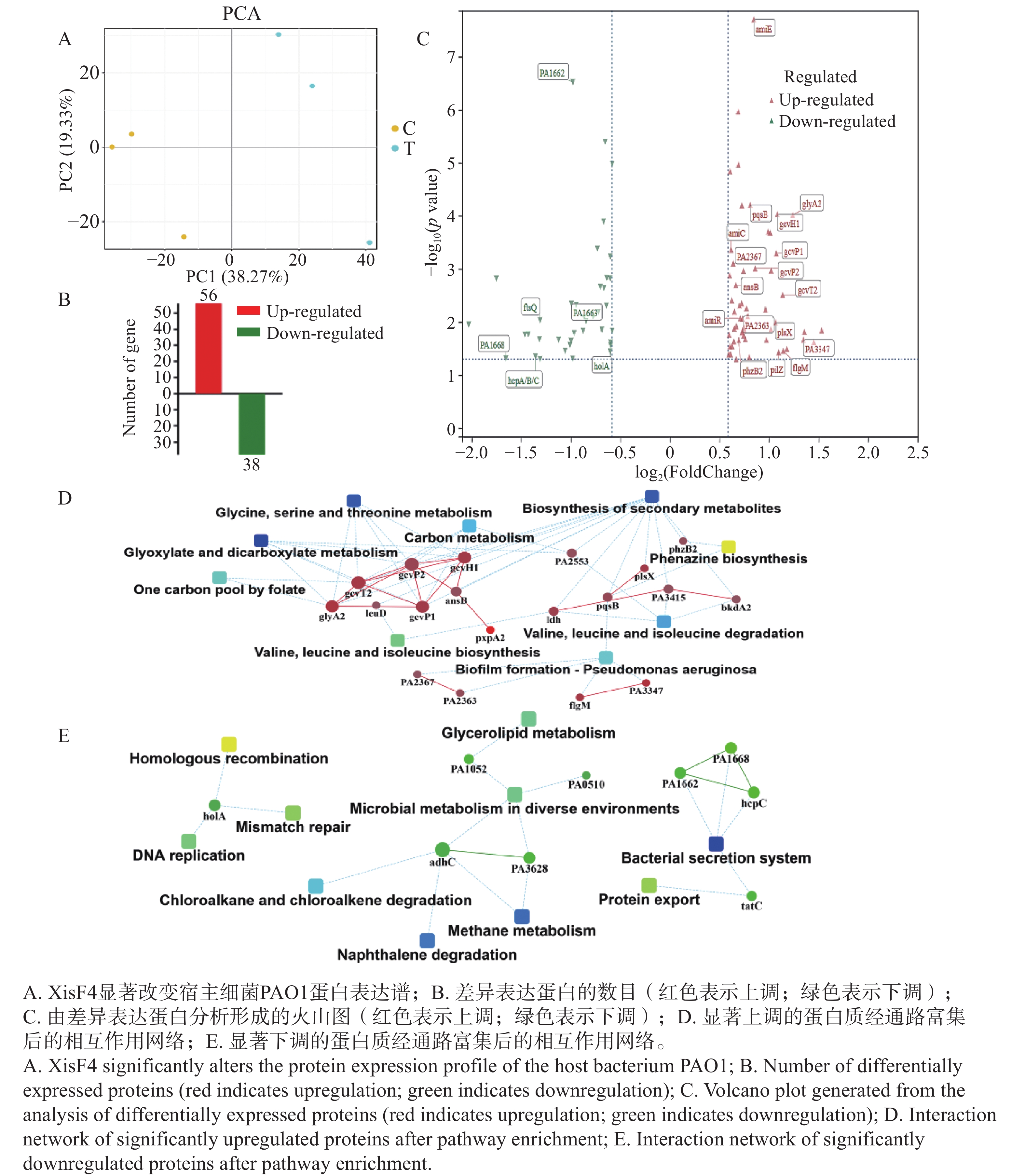
为明确XisF4的调控范围是否突破噬菌体基因并延伸至宿主细菌基因组,本研究将重组质粒pHERD20T::xisF4转化至铜绿假单胞菌PAO1的Pf4噬菌体敲除突变株(PAO1 ΔPf4)感受态细胞中,同时将pHERD20T空载体转化至PAO1 ΔPf4感受态细胞作为对照组。二者培养基中添加20 mmol·L−1阿拉伯糖作为表达诱导剂。诱导表达5 h后,提取细菌总蛋白并完成蛋白组测序。总蛋白鉴定数为3 571个,总肽段鉴定数为36 734个。经分析发现,XisF4显著改变了宿主细菌PAO1蛋白表达谱(图5-A)。经统计,共有94个蛋白差异表达(图5-B),其中56个蛋白显著上调,包括有病原菌致病相关蛋白如群体感应通路上的PqsB和PhzB2,四型菌毛蛋白PilZ,及甘氨酸代谢相关蛋白GcvP1,GcvP2,GcvH1,GcvT2和GlyA2(图5-C);38个显著下调蛋白包括细菌分泌系统蛋白HcpA/B/C,DNA复制蛋白HolA及参与细菌分裂的FtsQ(图5-C)。进一步对表达差异显著蛋白进行系统性的通路富集分析和蛋白相互作用网络构建,证实了甘氨酸/丝氨酸/苏氨酸代谢,生物被膜形成,绿脓素合成等相关通路受XisF4显著上调(图5-D),而蛋白分泌/外排,DNA错配修复/合成等相关的通路被显著抑制(图5-E)。综上总结为温和噬菌体编码的切离酶XisF4可显著影响宿主细菌多项不同生理功能蛋白的表达。
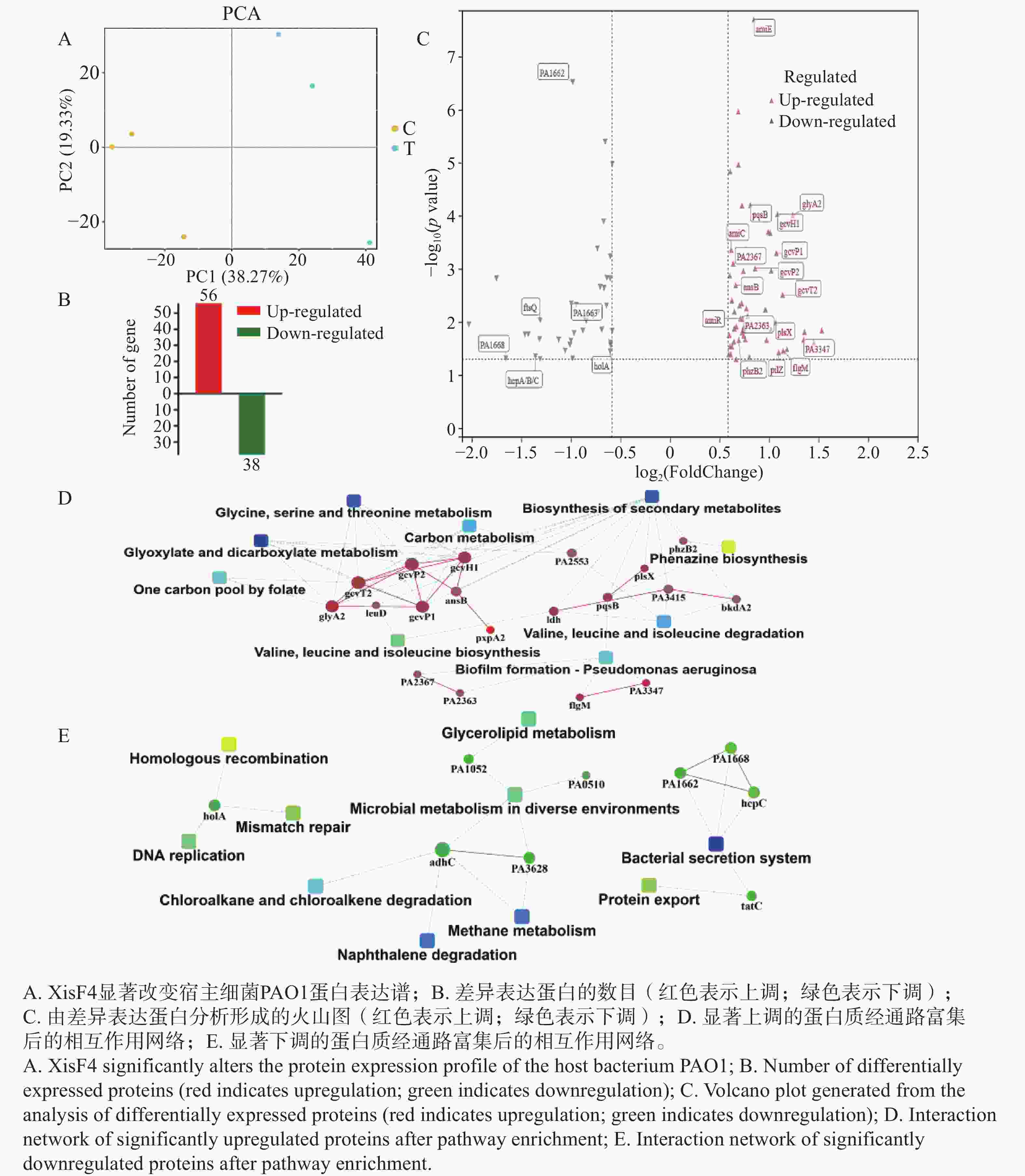
Figure 5. Impact of XisF4 on the proteomic profile of PAO1
-
通过RT-qPCR检测差异表达蛋白对应基因的mRNA水平变化,结果表明,在XisF4过表达条件下,自身编码基因xisF4的转录水平显著升高(图6-A),证实了诱导表达系统的有效性(图4-A)。并且,与蛋白质组学结果一致,群体感应与毒力相关基因pqsB和phzB2的转录水平分别上调了约3.2倍和3.4倍;但编码VI型分泌系统组分的hcpC基因转录水平则显著下调(图6-A)。这些数据在转录层面证实了XisF4能够调控宿主细菌的毒力与分泌系统相关基因的表达。
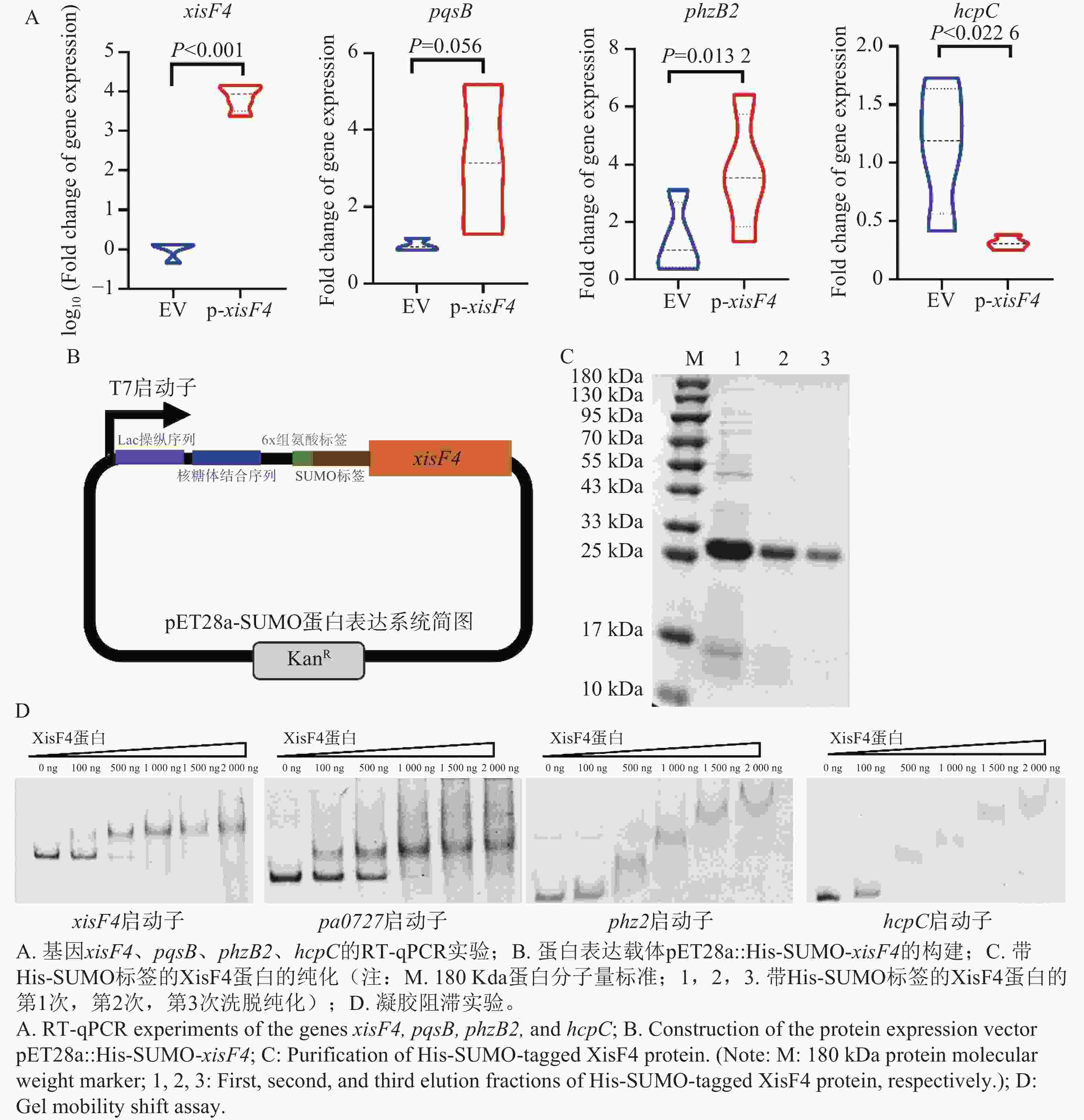
Figure 6. Verification of the bacterial host gene regulation by XisF4
构建pET28a::His-SUMO-xisF4蛋白表达载体(图6-B),并在大肠杆菌BL21(DE3)中表达,同时进一步纯化带有His-SUMO标签的重组XisF4蛋白,SDS-PAGE分析显示目的蛋白纯度较高(图6-C)。这说明XisF4是通过直接结合靶基因启动子区域来实施转录调控。
随后凝胶阻滞实验(EMSA)检测纯化的XisF4蛋白与潜在靶基因启动子片段的直接结合能力。实验选取了4个启动子片段,包括XisF4自身基因的启动子和噬菌体Pf4复制起始蛋白编码基因pa0727启动子作为阳性对照,及2个宿主基因的启动子:phz2(绿脓素合成操纵子)和hcpC(VI型分泌系统)。如图6-D所示,随着XisF4蛋白浓度的增加,xisF4和pa0727启动子DNA条带均出现明显的剂量依赖性迁移滞后现象,表明XisF4能与之直接结合,这与前期研究结果相符[13]。更重要的是,在相同实验条件下,phz2和hcpC启动子片段也表现出清晰的蛋白与DNA的复合物条带,且滞后程度随蛋白浓度增加而增强,因此在体外证实了XisF4能够直接结合到宿主基因phz2和hcpC的启动子区域。
-
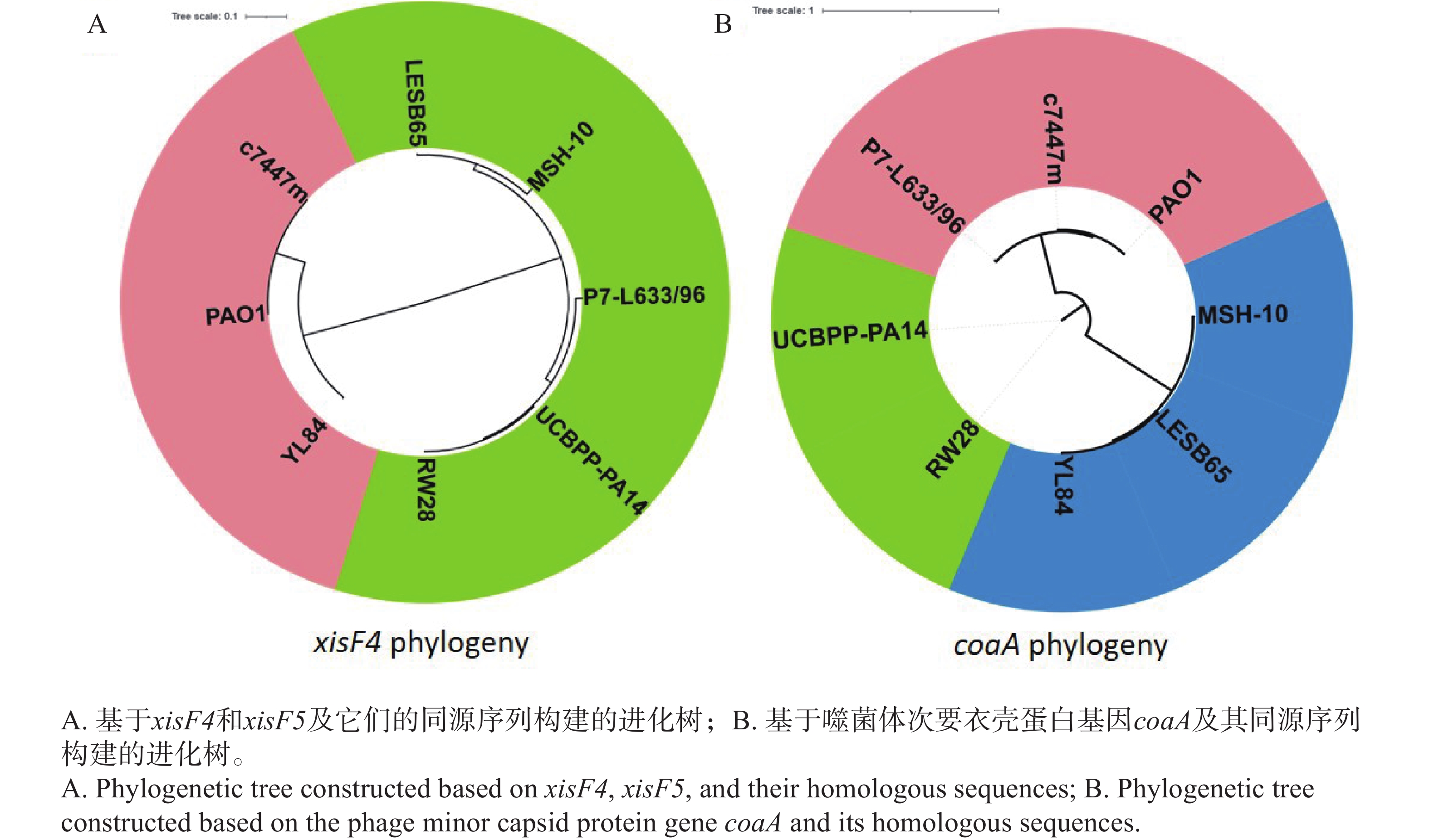
由于切离酶主要分为XisF4和XisF5两种类型[13],因此它们的基因编码序列及代表性的同源序列被调取并构建了系统发育树。侵染菌株PAO1、YL84、c7447m的丝状噬菌体编码同一类型切离酶(XisF4型),因而它们聚为一簇;而寄生于菌株LESB65、MSH-10、P7-L633/96、UCBPP-PA14、RW28的丝状噬菌体编码XisF5型切离酶(图7-A),则形成另一个进化分支。然而,这些丝状噬菌体在次要衣壳蛋白基因coaA的系统发育树上分布在多个不同分支:1)侵染菌株P7-L633/96且携带XisF5型切离酶的噬菌体与侵染菌株PAO1且携带XisF4型切离酶的噬菌体在coaA基因系统发育树上聚为一簇;2)携带XisF4型切离酶的噬菌体(YL84-Pf)与侵染菌株LESB65、MSH-10且携带XisF5型切离酶的噬菌体(LESB65-Pf和MSH-10-Pf)在coaA基因系统发育树上聚为另一簇(图7-B)。系统发育分析结果表明,虽然丝状噬菌体Pf基因组同时携带切离酶基因与衣壳蛋白基因,但它们在遗传进化规律上有显著差异。XisF4是一种多功能切离酶与转录调控因子,主要识别并结合噬菌体基因组中的特定DNA序列,在原噬菌体切离、复制启动等过程中发挥核心作用;次要衣壳蛋白CoaA主要参与噬菌体颗粒的结构组成,能识别并吸附宿主细菌的IV型菌毛,进而启动噬菌体感染。蛋白功能的不同可能是决定二者进化特征差异的主要原因。
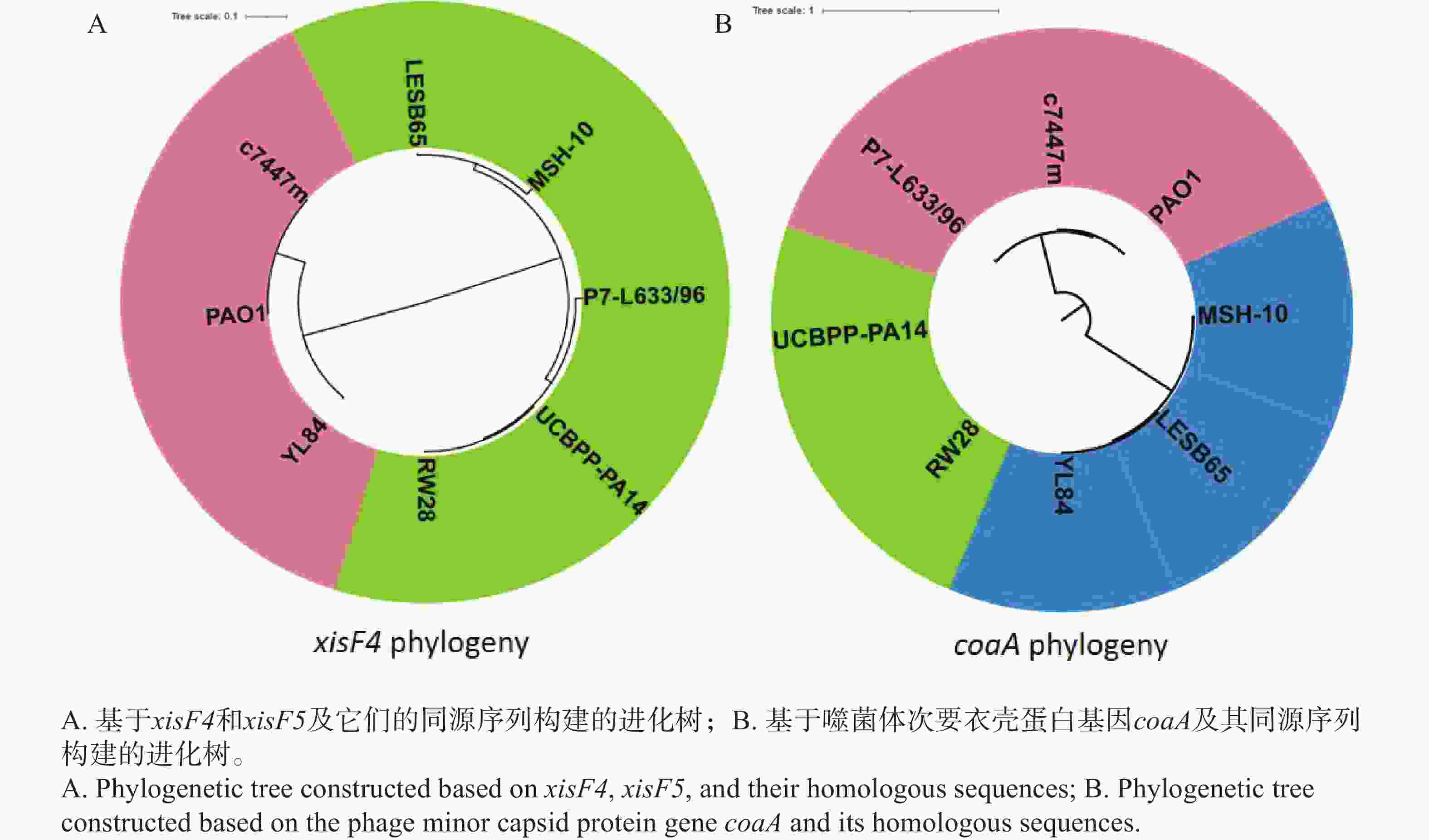
Figure 7. Phylogenetic analysis of XisF4 protein
-
铜绿假单胞菌作为临床上一种重要的机会性致病菌,其耐药性与持久感染能力与生物被膜形成密切相关。丝状噬菌体Pf是侵染铜绿假单胞菌的温和噬菌体[15],它在调控病原菌生物被膜形成过程中起到重要作用。在原噬菌体状态时,Pf编码的多个基因产物,如小分子RNA PhrD[16]和蛋白PfrT[17]等,能通过调控宿主细菌基因表达影响生物被膜;同时Pf还能通过编码的切离酶XisF4调控原噬菌体切离。游离状态下能与高分子聚合物形成纺锤状晶体,赋予细菌一层保护性屏障抵抗干燥和抗生素[18]。此外,丝状噬菌体Pf还能增强细菌感染致病力。当游离的噬菌体被胞吞进入上皮细胞或免疫细胞内,噬菌体转录的RNA会被胞内的病毒模式识别受体TLR3识别并激活I型干扰素通路,抑制NF-κB炎性信号通路,导致免疫细胞吞噬细菌的效率减弱,细菌逃逸免疫,造成小鼠存活率降低[19]。本研究通过生物信息学、蛋白组学与进化分析,系统揭示了XisF4的分子特性及其对宿主生理功能更广泛影响。本研究首先对XisF4蛋白进行了系统的生物信息学分析,结果表明,XisF4是一个小型不稳定蛋白,富含α螺旋与无规卷曲结构,缺乏跨膜域与信号肽,提示其为一胞内蛋白。更重要的是,本研究鉴定出其MerR型HTH结构域[20],并预测了其特异性结合的DNA序列模体,这与既往研究中XisF4与xisF4、pf4r、intF4及pilA启动子结合的结论一致[13-14],从结构基础上解释了其作为转录调控因子的作用机制。大多数MerR型转录调节因子能够响应环境刺激,如重金属、氧化应激或抗生素[21]。这也解释了为什么一些环境应激压力信号,如氧化压力、DNA损伤、生物被膜形成后期等,会显著激活XisF4的生物活性,进而导致原噬菌体Pf4的切离,环化复制,最终释放大量的噬菌体颗粒[22]。
进一步的蛋白组学分析发现,XisF4的表达显著改变了94个宿主蛋白的表达水平,其中56个上调、38个下调。一方面,多个与致病性密切相关的蛋白如PqsB、PhzB2、PilZ等显著上调,提示XisF4可能通过增强群体感应系统及菌毛合成;此外甘氨酸代谢通路相关蛋白(GcvP1/P2、GcvH1、GcvT2、GlyA2)的上调,可能与噬菌体整合位点位于甘氨酸tRNA基因附近有关[23],体现了原噬菌体与宿主代谢网络的深度互作。另一方面,XisF4下调了包括VI型分泌系统组分(HcpA/B/C)、DNA复制蛋白HolA及细胞分裂蛋白FtsQ在内的多个关键功能蛋白,这表明XisF4可能抑制宿主分泌效应蛋白的能力、影响基因组稳定性与细胞分裂,从而在感染过程中调控宿主应激反应与生存策略。进化分析结果显示,XisF4与XisF5分属2个不同的进化分支,且与衣壳蛋白CoaA的进化轨迹不一致,这说明丝状噬菌体在不同菌株中通过水平基因转移与模块化重组获得不同的切离酶与结构蛋白组合。这种遗传灵活性可能是丝状噬菌体适应不同宿主环境的重要机制。由于噬菌体结构蛋白基因CoaA与噬菌体的感染性和宿主识别密切相关,因此需要不同结构蛋白来与不同细菌宿主表面受体相结合。噬菌体结构蛋白的进化树可能更多地反映了衣壳蛋白对特定宿主的适应过程。综上所述,XisF4作为Pf4噬菌体的切离酶不仅负责原噬菌体的切离,更可以作为一个多效转录调控因子,广泛影响宿主细菌的毒力、代谢、分泌系统及分裂过程。本研究从分子结构、功能调控及进化角度深化了对XisF4的认识,为后续针对铜绿假单胞菌噬菌体—宿主互作机制的相关干预策略研发提供了理论依据。
Bioinformatics characterization, host proteome regulatory, and phylogenetic evolution analysis of the filamentous phage Pf4 excisionase XisF4
DOI: 10.15886/j.cnki.rdswxb.20250198
- Received Date: 2025-12-17
- Accepted Date: 2026-02-09
- Rev Recd Date: 2026-01-29
-
Key words:
- Pseudomonas aeruginosa /
- filamentous bacteriophage /
- excisionase /
- bioinformatic analysis /
- proteomics analysis
Abstract: The excisionase XisF4, encoded by the filamentous bacteriophage Pf4, is a key protein mediating prophage excision and transcriptional regulation. The molecular characteristics and biological functions of the excisionase XisF4 were systematically elucidated through the integration of bioinformatics, proteomics, and evolutionary analyses. Bioinformatics analysis revealed that XisF4 consists of 71 amino acids and contains a typical MerR-type helix-turn-helix DNA-binding domain. Induced expression and proteomic analysis showed that XisF4 expression led to the differential expression of 94 host proteins, significantly enriched in pathways such as glycine metabolism, biofilm formation, and pyocyanin biosynthesis, while simultaneously inhibiting the expression of proteins associated with the type VI secretion system (e.g., HcpC) and DNA replication. RT-qPCR validated transcriptional changes in key genes, and further EMSA experiments confirmed that purified XisF4 protein directly binds to the promoter regions of host genes phz2 and hcpC, demonstrating its direct transcriptional regulatory capability. Phylogenetic analysis revealed that XisF4 and XisF5 belong to distinct evolutionary branches, and that their evolutionary trajectories differ from that of the bacteriophage capsid protein CoaA, suggesting that independent genetic events may have occurred between the regulatory and structural modules of filamentous bacteriophages during evolution. All the results demonstrate that XisF4 is not only a key enzyme for bacteriophage excision but also functions as a pleiotropic transcriptional regulator, extensively involved in regulating virulence, metabolism, and stress adaptation in the host Pseudomonas aeruginosa. These findings provide an important theoretical foundation for a deeper understanding of bacteriophage-host interaction mechanisms and the development of potential intervention strategies.
| Citation: | Zhang Jiawen, Shi Shengmei, Wen Zewen, Li Peiyu, Yu Zhijian, Yang Qiong, Li Shuo, Tang Mingxing, Wei Lu. Bioinformatics characterization, host proteome regulatory, and phylogenetic evolution analysis of the filamentous phage Pf4 excisionase XisF4[J]. Journal of Tropical Biology. doi: 10.15886/j.cnki.rdswxb.20250198

|
 DownLoad:
DownLoad: