-
珊瑚礁生态系统是重要的热带海洋生态系统之一,因其具有较高的生物多样性而被誉为“海洋中的热带雨林”[1]。长棘海星(Acanthaster spp.)喜食珊瑚礁生态系统的框架生物造礁珊瑚。长棘海星群体暴发被认为是近年来全球珊瑚礁生态系统退化的重要原因之一[2-3]。 2009 年长棘海星群体暴发导致西沙群岛部分礁区的活珊瑚覆盖率降至不足1%[4-5],珊瑚生物多样性显著降低[6]。此外,遭受长棘海星啃食后留下的珊瑚骨骼会因生物侵蚀和海水运动而塌陷, 从而降低珊瑚礁结构的复杂性,并致使礁栖生物的种类和密度及生物多样性下降 [7-9]。
礁栖鱼类可以捕食不同生长阶段的长棘海星,进而能抑制长棘海星群体暴发[10]。用于解释长棘海星群体暴发机理的“捕食者移除假说”认为,当长棘海星捕食者大幅减少时,长棘海星群体将可能因上行控制降低而发生暴发[11]。Cowan 等[12]研究发现,常见的雀鲷科和蝴蝶鱼科鱼类会主动地摄食长棘海星的配子和幼虫,鲀形目的纹腹叉鼻鲀(Arothron hispidus)等鱼类可直接捕食长棘海星成体[13–14]。这些鱼类捕食者在控制长棘海星群体数量和种群动态等方面发挥着重要作用[15]。近年来有研究者运用环境DNA 技术来探究长棘海星捕食者的种类和数量。如Kroon 等[11]通过ddPCR 技术和特异性引物[16](F:5'-CCGGACGCCTATAC-3';R:5'-AGTGGTTCGCTGGGAAGTGAAGG-3')检测了澳大利亚大堡礁海洋公园鱼类肠道内容物中的长棘海星线粒体细胞色素C 氧化酶亚基Ⅰ(mitochondrial cytochrome oxidase subunit Ⅰ , CoTS-mtCOⅠ)基因片段,发现豹纹鳃棘鲈(Plectropomus leopardus)、星斑裸颊鲷(Lethrinus nebulosus)、宅泥鱼(Dascyllus aruanus)、黑斑盘雀鲷(Dischistodus melanotus)4 种长棘海星潜在鱼类捕食者。张颖等[17]采用PCR 技术检测了鱼肠道内容物中的长棘海星DNA,发现了颊吻鼻鱼(Naso lituratus)、网纹宅泥鱼(Dascyllus reticulatus)、三叶唇鱼(Cheilinus trilobatus)和赤鳍裸颊鲷(Lethrinus erythropterus)4 种长棘海星潜在鱼类捕食者。尽管“捕食者移除假说”难以解释全球发生的所有长棘海星群体暴发事件[13, 18],但对长棘海星的鱼类捕食者种类进行全面了解将有利于控制长棘海星群体数量,减少长棘海星暴发风险,并提高珊瑚礁生态系统的自然恢复力[19]。
中国南海礁区自2020年开始新一轮的长棘海星(Acanthaster cf. solaris) 群体暴发。为解析中国南海长棘海星的群体暴发机理和防控长棘海星群体暴发提供理论依据与科学指导,本研究根据形态学特征与18S 分子标签鉴定了在西沙礁区 23 个站点采捕的鱼类,然后采用PCR 技术检测了其肠道内容物 DNA 中的 CoTS-mtCOⅠ 基因片段,进而分析中国西沙礁区长棘海星的潜在鱼类捕食者及其摄食能力。
-
2023 年 4 月8~26日在 23 个西沙海域站点(表1)网捕鱼类,将全部渔获置于−20 ℃冰箱暂存。待渔获运抵实验室后,根据 FishBase(
http://fishbase. org/ )的形态学特征鉴定样品的所属物种。首先从每种鱼类中选取 1 尾样品,取米粒大的肌肉组织样置于含有 500 mL DNA lysis buffer (10 mmol·L−1 Tris-HCl (pH 8.0),0.5% SDS,100 mmol·L−1 EDTA,200 µg·mL−1 Protease K)的离心管中,−80 ℃冰箱中保存至 DNA 提取,用于进一步的样品鉴定。然后对每个站点中的每种鱼类随机挑选 3 尾(不足 3 尾时取全部),参考 Kroon 等[11]的方法取样肠道内容物,即解剖鱼体后取出肠道,在前端剪取 1~3 cm 长的肠道,置于装有2 mL DNA lysis buffer 的离心管中,−80 ℃保存,用于后续的 DNA 提取。站点Station 邻近岛礁
nearby reefs经度(E)longitude 纬度(N)
latitudeS1 北礁Beijiao 111° 26.3168 ′17°02.0614′ S2 西沙洲Xishazhou 112° 10.1718 ′16° 56.1076 ′S3 南沙洲Nanshazhou 112° 24.7123 ′16° 56.5063 ′S4 永兴岛Yongxingdao 112° 17.7411 ′16° 47.5152 ′S5 东岛Dongdao 112° 54.3976 ′16° 47.0225 ′S6 银砾滩Yinlitan 112° 13.0569 ′16° 42.5217 ′S7 高尖石Gaojianshi 112° 41.5052 ′16° 34.8471 ′S8 石屿Shiyu 111° 50.8642 ′16° 30.3492 ′S9 珊瑚岛Shanhudao 111° 35.1539 ′16° 34.9625 ′S10 金银岛Jinyindao 111° 28.3318 ′16° 27.8815 ′S11 华光礁Huaguangjiao 111° 35.2902 ′16° 15.8143 ′S12 华光礁Huaguangjiao 111° 32.5180 ′16°06.1935′ S13 华光礁Huaguangjiao 111° 53.4598 ′16° 11.3570 ′S14 盘石屿Panshiyu 111° 52.1960 ′16°02.3941′ S15 滨湄滩Binmeitan 112° 36.4320 ′16° 16.6061 ′S16 浪花礁Langhuajiao 112° 29.8840 ′16°06.4386′ S17 玉琢礁Yuzhuojiao 112°08.8745′ 16° 20.3769 ′S18 玉琢礁Yuzhuojiao 112°08.4118′ 16°05.5609′ S19 湛涵滩Zhanhantan 112° 50.7861 ′16° 20.2253 ′S20 永兴岛Yongxingdao 112° 20.8649 ′16° 36.2650 ′S21 石屿Shiyu 111° 58.6927 ′16° 30.6776 ′S22 西沙洲Xishazhou 111° 57.7774 ′17°00.3571′ S23 盘石屿Panshiyu 111° 38.5111 ′16°01.0240′ Table 1. Sampling sites and latitude/longitude
-
采用 18S 分子标签进一步鉴定鱼类物种。首先,取出冻存的肌肉样品,解冻后依次加入 500 μL 裂解液(10 mmol·L−1 Tris-HCl (pH=8.0),10 mmol·L−1 EDTA(pH=8.0),10 mmol·L−1 NaCl,1% 2-巯基乙醇,10 μL 蛋白酶k(10 mg·mL−1)、65 μL 的 10%SDS),56 ℃温浴 1 h ,每隔 15 min 颠倒摇动数次,使样品充分裂解。室温下 8 000 g 离心 3 min ,吸取 500 μL 上清液,加入500 μL的V苯酚:V氯仿:V异戊醇=25:24:1混合液。颠倒混匀后,室温13 000 g 离心 10 min ,吸取400 μL 上清液,加入400 μL的V氯仿:V异戊醇=24:1混合液。再次颠倒混匀后,室温13 000 g 离心 10 min ,吸取 200 μL上清液至新的离心管中,加入 400 μL无水乙醇。混匀后静置15 min ,室温3 000 g 离心 10 min ,倒去上清液,加入 1 mL 70%乙醇。室温13 000 g 离心 5 min ,倒去上清液后,室温干燥15 min。最后加入100 μL TE 溶解 DNA 沉淀,用于后续的 PCR实验。
以提取的肌肉组织DNA 为模板,采用18S 通用引物对(F : 5'-GCCAGTAGTCATATGCTTGTCT-3' ;R:5'-GGAGCTGGAATTACCGC-3')PCR 扩增 18S 分子标签序列。扩增体系:2.5 μL 10×PCR Buffer,2 μL dNTP(各 2.5 mmol·L−1),1 μL 引物 F(10 μmol·L−1),1 μL 引物R(10 μmol·L−1),1 μL DNA 模板,0.15 μL rTaq,17.35 μL DEPC 水。扩增条件为95 ℃ 5 min;95 ℃ 30 s,55 ℃ 30 s,72 ℃ 30 s,35 个循环;72 ℃ 10 min,4 ℃保存。PCR 产物经琼脂糖凝胶电泳确定为特异性扩增后,送至生工生物工程(上海)股份有限公司进行核酸测序。将获得的 18S 序列(表2)比对至NCBI 的 nt 数据库,根据序列同源性进一步确定样品的所属物种[20]。
序号ID 种名
Species分类地位
Taxonomy18S序列号
18S Accession Number1 珠蝴蝶鱼 Chaetodon kleinii 刺尾鱼目 Aanthuriformes 蝴蝶鱼科 Chaetodontidae EF616825.1 2 三间火箭 Chelmon rostratus 刺尾鱼目 Acanthuriformes 蝴蝶鱼科 Chaetodontidae EF616876.1 3 弓月蝴蝶鱼 Chaetodon lunulatus 刺尾鱼目 Acanthuriformes 蝴蝶鱼科 Chaetodontidae OP035277.1 4 角镰鱼 Zanclus cornutus 刺尾鱼目 Acanthuriformes 镰刀鱼科 Zanclidae NC_009852.1 5 黄斑蓝子鱼 Siganus canaliculatus 刺尾鱼目 Acanthuriformes 篮子鱼科 Siganidae AF055600.1 6 银蓝子鱼 Siganus argenteus 刺尾鱼目 Acanthuriformes 篮子鱼科 Siganidae PP140670.1 7 少鳞燕鳐 Cypselurus oligolepis 颌针鱼目 Beloniformes 飞鱼科 Exocoetidae LC458117.1 8 黑背圆颌针鱼 Tylosurus melanotus 颌针鱼目 Beloniformes 颌针鱼科 Belonidae MH248221.1 9 青翼须唇飞鱼 Cheilopogon cyanopterus 颌针鱼目 Beloniformes 飞鱼科 Exocoetidae NC_036721.1 10 小鳍针飞鱼 Oxyporhamphus micropterus 颌针鱼目 Beloniformes 鱵科 Hemiramphidae LC715466.1 11 花鳍燕鳐 Cypselurus poecilopterus 颌针鱼目 Beloniformes 飞鱼科 Exocoetidae OP035088.1 12 六带鲹 Caranx sexfasciatus 鲹形目 Carangiformes 鲹科 Carangidae OP035090.1 13 脂眼凹肩鲹 Selar crumenophthalmus 鲹形目 Carangiformes 鲹科 Carangidae NC_023954.1 14 长颌似鲹 Scomberoides commersonnianus 鲹形目 Carangiformes 鲹科 Carangidae LC672493.1 15 细鳞圆鲹 Decapterus macarellus 鲹形目 Carangiformes 鲹科 Carangidae NC_026718.1 16 单棘豹鲂鮄 Daicocus peterseni 豹鲂鮄目 Dactylopteriformes 豹鲂鮄科 Dactylopteridae NC_003194.1 17 无斑拟羊鱼 Mulloidichthys vanicolensis 羊鱼目 Mulliformes 羊鱼科 Mullidae NC_024188.1 18 圆口副绯鲤 Parupeneus cyclostomus 羊鱼目 Mulliformes 羊鱼科 Mullidae PP140669.1 19 条斑副绯鲤 Parupeneus barberinus 羊鱼目 Mulliformes 羊鱼科 Mullidae NC_082530.1 20 奈氏眶灯鱼 Diaphus knappi 灯笼鱼目 Myctophiformes 灯笼鱼科 Myctophidae LC146181.1 21 犬牙锥齿鲷 Pentapodus caninus 鲈形目 Perciformes 金线鱼科 Nemipteridae LC036827.1 22 红裸颊鲷 Lethrinus haematopterus 鲈形目 Perciformes 裸颊鲷科 Lethrinidae LC640073.1 23 黑带鳞鳍梅鲷 Pterocaesio tile 鲈形目 Perciformes 乌尾鮗科 Caesionidae NC_004408.1 24 宝石大眼鲷 Priacanthus hamrur 鲈形目 Perciformes 大眼鲷科 Priacanthidae LC649119.1 25 灰鳍异大眼鲷 Heteropriacanthus cruentatus 鲈形目 Perciformes 大眼鲷科 Priacanthidae OP035148.1 26 白边侧牙鲈 Variola albimarginata 鲈形目 Perciformes 鮨科 Serranidae NC_022139.1 27 六带线纹鱼 Grammistes sexlineatus 鲈形目 Perciformes 线纹鱼科 Grammistidae AP006794.1 28 东方尖唇鱼 Oxycheilinus orientalis 鲈形目 Perciformes 隆头鱼科 Labridae LC104664.1 29 蜂巢石斑鱼 Epinephelus merra 鲈形目 Perciformes 石斑鱼科 Epinephelidae NC_022509.1 30 鲔 Euthynnus affinis 鲭形目 Scombriformes 鲭科 Scombridae NC_025934.1 31 鲣 Katsuwonus pelamis 鲭形目 Scombriformes 鲭科 Scombridae KM605252.1 32 黄鳍金枪鱼 Thunnus albacares 鲭形目 Scombriformes 鲭科 Scombridae NC_014061.1 33 沙氏刺鲅 Acanthocybium solandri 鲭形目 Scombriformes 鲭科 Scombridae NC_067731.1 34 圆舵鲣 Auxis rochei 鲭形目 Scombriformes 鲭科 Scombridae MK548578.1 35 蛇鲭 Gempylus serpens 鲭形目 Scombriformes 蛇鲭科 Gempylidae OP035147.1 36 扁舵鲣 Auxis thazard 鲭形目 Scombriformes 鲭科 Scombridae MK801690.1 37 棘鲯鳅 Coryphaena equiselis 鲹形目 Carangiformes 鲯鳅科 Coryphaenidae MH576916.1 38 琉璃玉鲳 Psenes cyanophrys 鲭形目 Scombriformes 圆鲳科Nomeidae NC_082747.1 39 鳞首方头鲳 Cubiceps squamiceps 鲭形目 Scombriformes 圆鲳科 Nomeidae PQ492202.1 40 细斑前孔鲀 Cantherhines pardalis 鲀形目 Tetraodontiformes 单棘鲀科 Monacanthidae OR499746.1 41 兔头鲀 Lagocephalus 鲀形目 Tetraodontiformes 四齿鲀科
TetraodontidaeLC738765.1 42 暗鳍兔头鲀 Lagocephalu gloveri 鲀形目 Tetraodontiformes 四齿鲀科
TetraodontidaeNC_059716.1 Table 2. Fish species, taxonomic status and 18S sequence numbers
-
采用 CTAB 法提取鱼类肠道内容物中的总DNA[21] ,通过琼脂糖凝胶电泳法和 Nanodrop 微量分光光度计分别检测鱼类肠道内容物总DNA的质量和浓度。使用长棘海星 CoTS-mtCOⅠ 片段的特异性扩增引物对(F:5'-TCCGACTACCCGGACGCCTATAC-3' ;R:5'- AGTGGTTCGCTGGGAAGTGAAGG-3'),以鱼类肠道内容物总 DNA作为模板,进行 PCR 扩增。扩增体系:2.5 μL 10×PCR Buffer 、2 μL dNTP(各 2.5 mmol·L−1)、1 μL 引物 F(10 μmol·L−1)、1 μL 引物R(10 μmol·L−1)、1 μL DNA 模板、0.15 μL rTaq,17.35 μL DEPC 水。扩增条件 95 ℃ 5 min;95 ℃ 30 s,60 ℃ 30 s,72 ℃ 30 s,35 个循环;72 ℃ 10 min,4 ℃保存。同时,提取长棘海星腕中部皮下肌肉的 DNA 并扩增 CoTS-mtCOⅠ 基因片段作为阳性对照组。
-
根据18S 分子标签序列和邻接法 (Neighbor joining),采用MEGA5.0 软件构建鱼类物种的分子系统进化树, 并经 1 000 次重复抽样(Bootstraps)检测其置信度。计算每种鱼类肠道内容物样品中的CoTS-mtCOⅠ 基因片段检出率,并采用 Origin 8.0 软件绘制圆锥图。采用卡方检验分析不同鱼类肠道内容物中 CoTS-mtCOⅠ 基因片段检出率的差异。以P<0.05 代表差异具有统计学意义。
CoTS-mtCOⅠ 基因片段检出率 = CoTS-mtCOⅠ 检出次数/检测样品总数
-
鉴定结果表明,所获鱼类隶属于 1 纲 9 目 24 科 37 属 42 种(表2)。形态学与 18S 分子标签二种方法鉴定出的鱼类物种基本一致,仅有一种经形态学鉴定为黑背圆颌针鱼(Tylosurus melanotus),而经18S 鉴定为鳄形圆颌针鱼 (T. crocodilus)。根据已有研究[22] 分析,黑背圆颌针鱼为鳄形圆颌针鱼的亚种,本研究将此鱼归为鳄形圆颌针鱼。在所有捕获鱼类中,细鳞圆鲹(Decapterus macarellus)渔获数量最多,且在大部分站点均有捕获;最少的种类为犬牙锥齿鲷(Pentapodus caninus),仅在华光礁捕获 1 条。
-
本研究通过从鱼肠道内容物总 DNA 中 PCR 扩增 CoTS-mtCOⅠ 基因片段来鉴定西沙礁区长棘海星的潜在鱼类捕食者。在无模板对照和单引物对照中,未见明显条带。在阳性对照中检测到单一条带,表明 CoTS-mtCOⅠ 基因片段的特异性扩增。进一步检测发现,在 6 种鱼类的肠道内容物 DNA 中存在 CoTS-mtCOⅠ 基因片段的阳性扩增,分别是珠蝴蝶鱼(Chaetodon kleinii)、三间火箭(Chelmon rostratus)、弓月蝴蝶鱼(Chaetodon lunulatus)、红裸颊鲷(Lethrinus haematopterus)、东方尖唇鱼(Oxycheilinus orientalis)和蜂巢石斑鱼(Epinephelus merra)(图1)。

Figure 1. CoTS-mtCOⅠ gene fragment amplification for total DNA of intestinal contents in six fish species and agarose gel electrophoresis
这 6 种鱼主要采集于西沙洲、永兴岛、石屿和金银岛附近礁区。其中,珠蝴蝶鱼采集于西沙洲附近礁区;三间火箭采集于西沙洲、永兴岛、石屿和金银岛附近礁区;弓月蝴蝶鱼采集于石屿附近礁区;红裸颊鲷采集于永兴岛和石屿附近礁区;东方尖唇鱼采自西沙洲附近海域;蜂巢石斑鱼源自西沙洲和金银岛附近礁区。
-
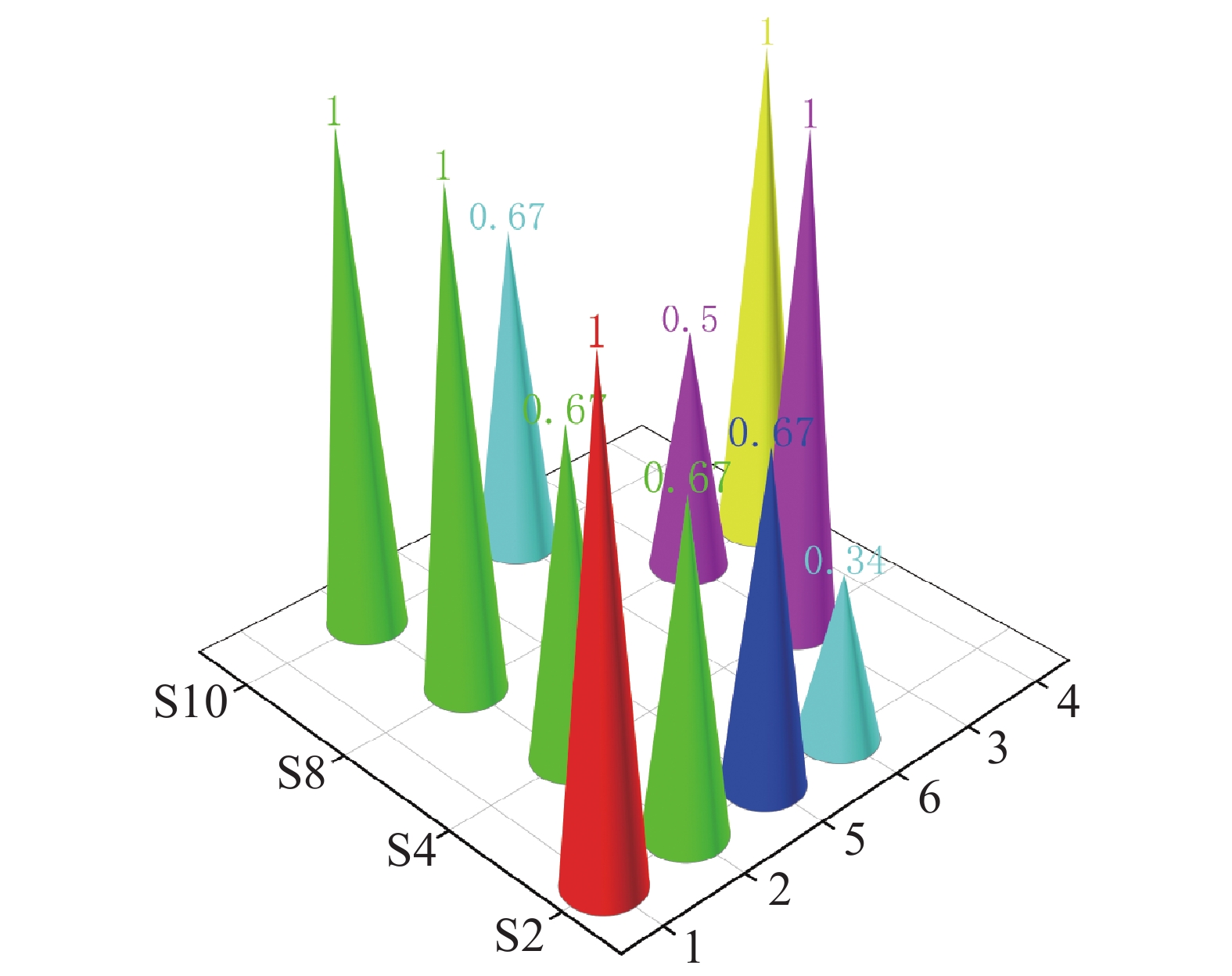
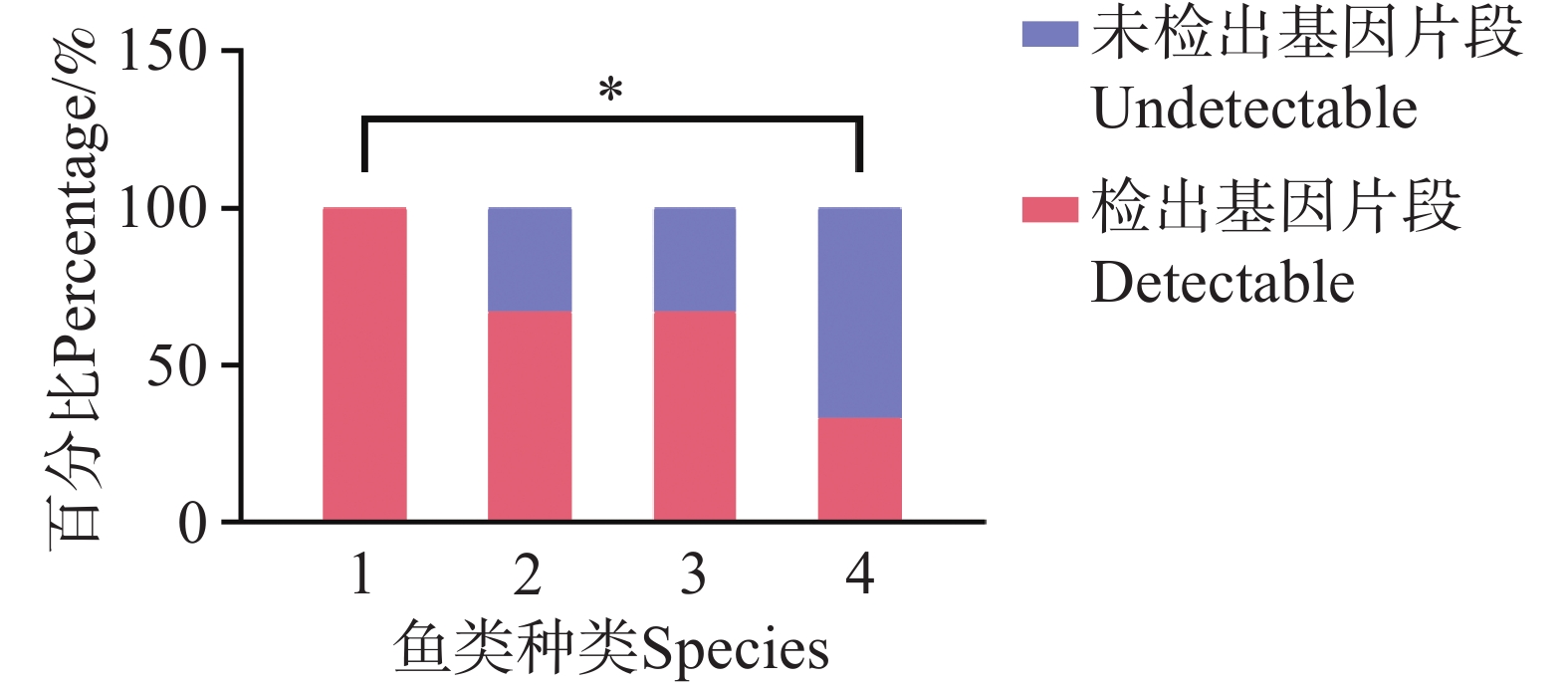
在珠蝴蝶鱼、弓月蝴蝶鱼的肠道内容物 DNA 中均检测到 CoTS-mtCOⅠ 基因片段。三间火箭和弓月蝴蝶鱼的 CoTS-mtCOⅠ 基因片段检出率在各个站点均大于 50%,而蜂巢石斑鱼和红裸颊鲷的检出率较低,分别为 33.4%和 50%(图2)。在西沙洲,珠蝴蝶鱼的检出率显著高于蜂巢石斑鱼(P<0.05)(图3)。在石岛、晋卿岛和金银岛,这些鱼类的检出率之间无显著差异。
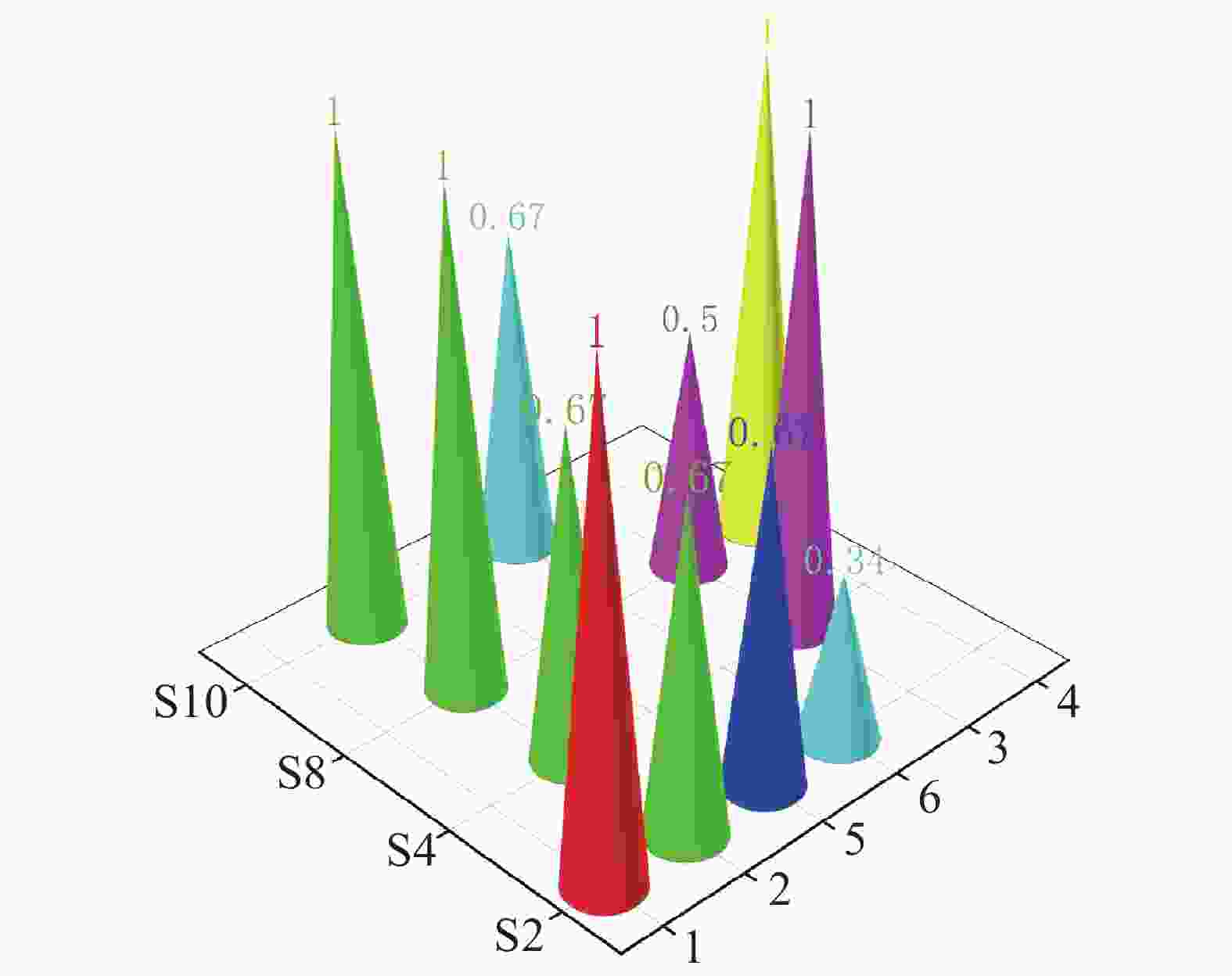
Figure 2. The detection rate of the CoTS-mtCOⅠ gene fragment in the intestinal contents of six fish species at four stations
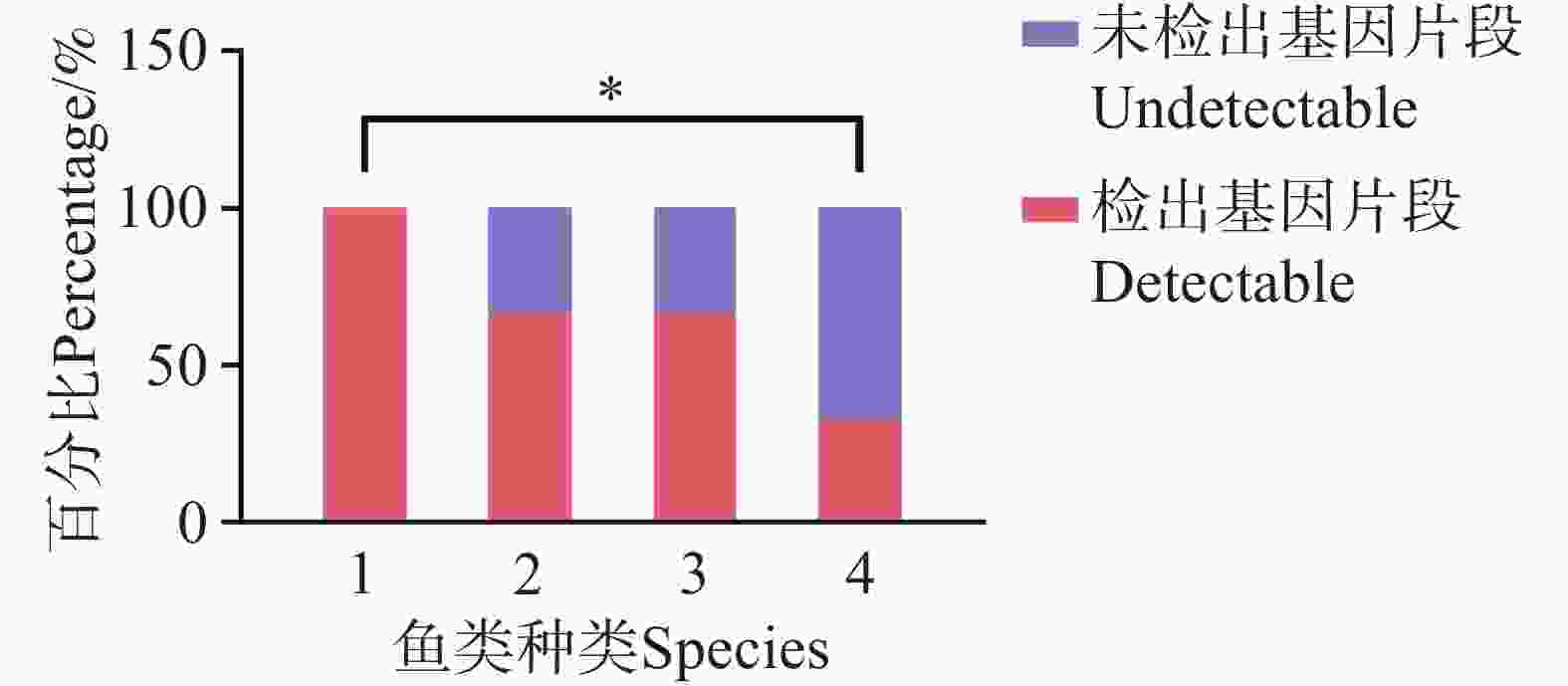
Figure 3. Detection rate analysis of CoTS-mtCOⅠ gene fragment in fish intestinal contents at Xishazhou station.
-
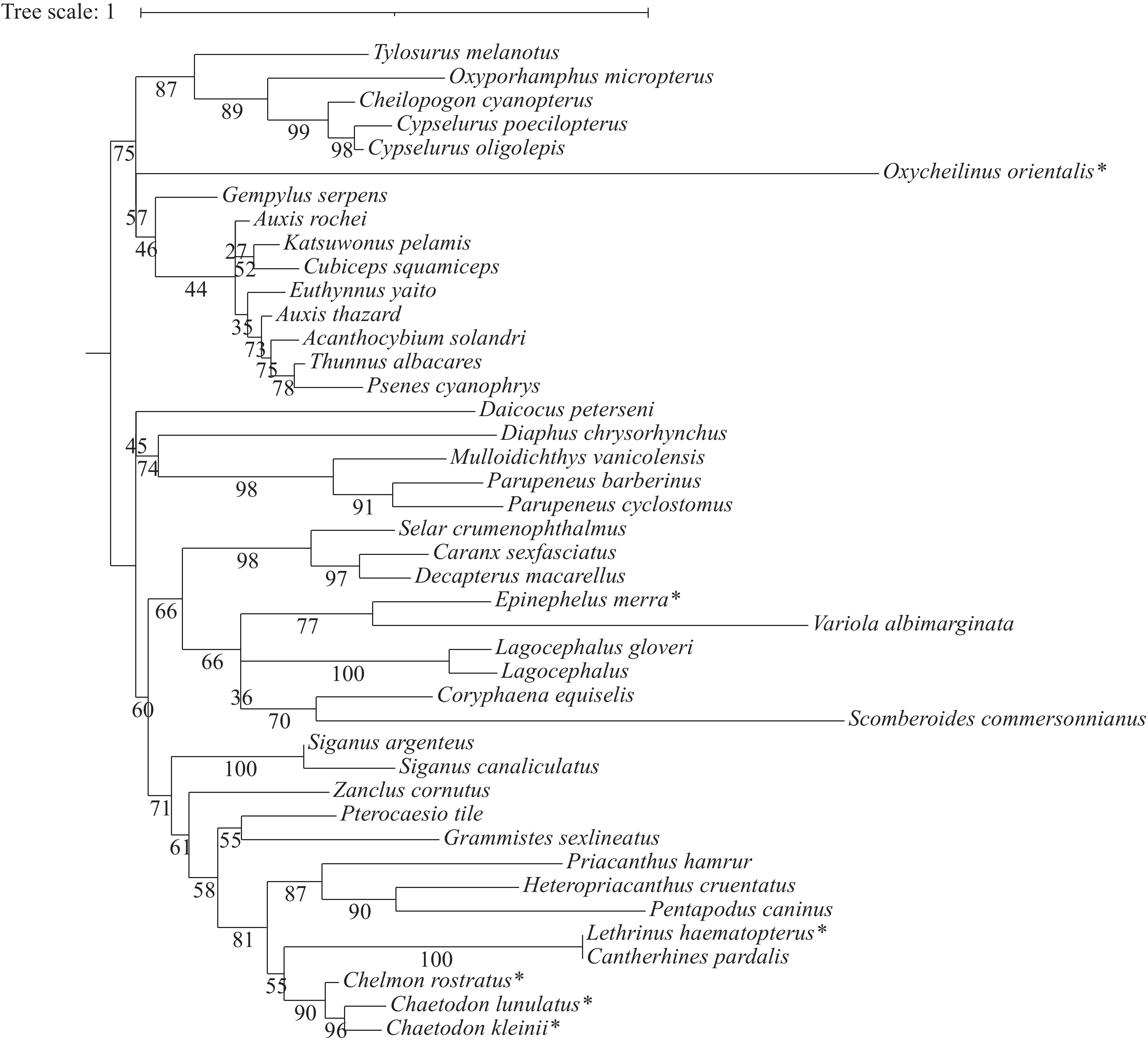
根据 42 种捕获鱼类的 18S 基因片段序列,利用邻接法构建系统发育树(图4)。在该进化树中,所有的同科鱼类均能汇聚为一支,如蝴蝶鱼科、飞鱼科、鯵科、羊鱼科和鲭科等。珠蝴蝶鱼、三间火箭与弓月蝴蝶鱼先聚到蝴蝶鱼科亚支,再归于刺尾鱼目分支。红裸颊鲷、东方尖唇鱼与蜂巢石斑鱼最终聚到鲈形目分支。
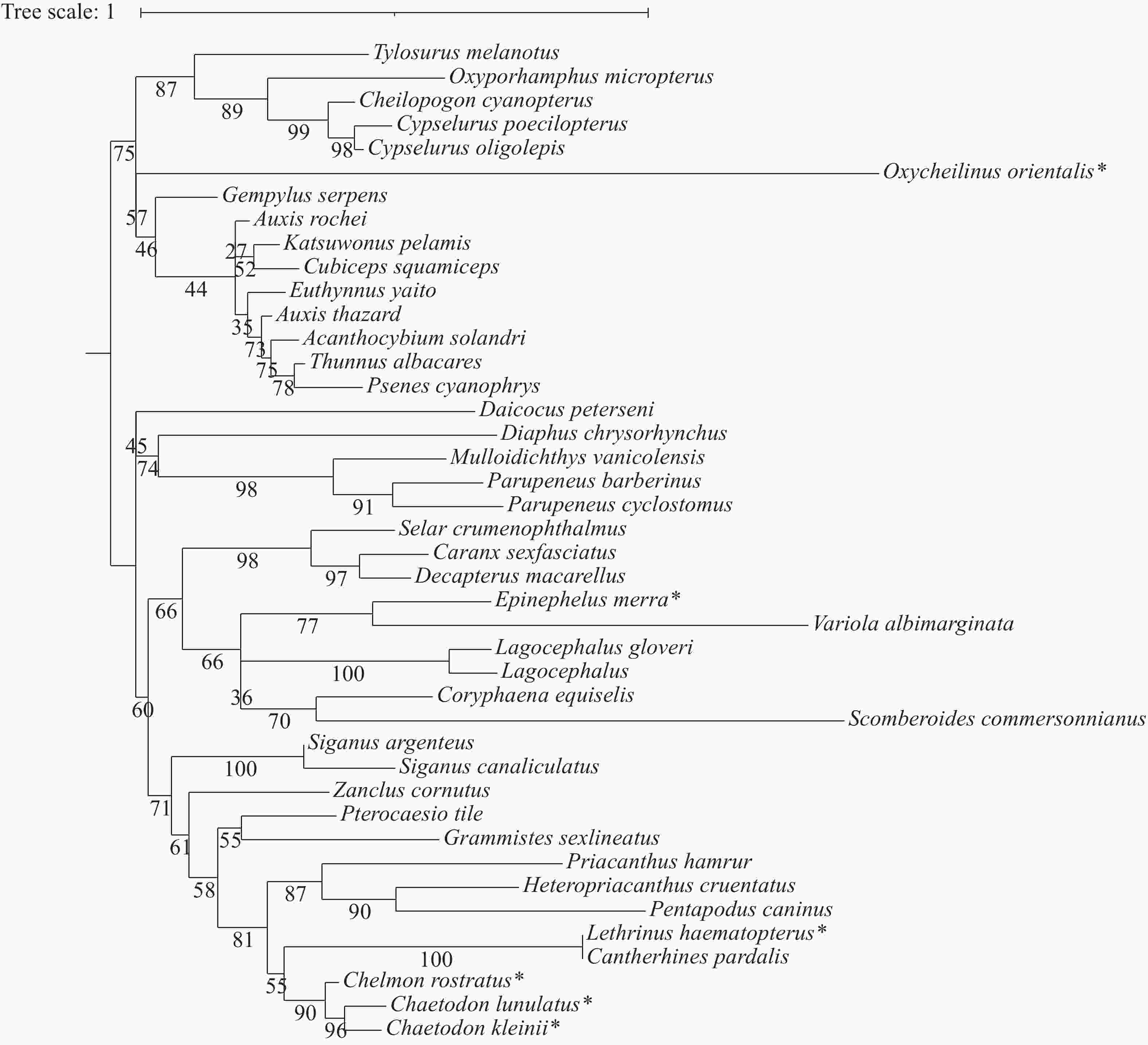
Figure 4. Molecular phylogenetic trees of 42 fish from Xisha island based on 18S gene fragment sequences constructed using Neighbor-Joining method
-
长棘海星群体暴发已导致珊瑚礁的生态系统退化及生物多样性下降,鱼类捕食者可控制长棘海星群体数量,从而降低长棘海星群体暴发频率及破坏程度[23]。因此,查明长棘海星的鱼类捕食者对珊瑚礁生态系统的保护很重要。本研究选择正遭受长棘海星暴发的西沙群岛礁区调查长棘海星的潜在鱼类捕食者物种及其摄食能力,以期更好地防控南海长棘海星暴发进而保护珊瑚礁生态系统。
至今已发现的长棘海星捕食者种类已达到132种,包括珊瑚礁鱼类和底栖无脊椎动物等[11, 14, 24-26] ,它们可摄食长棘海星配子、幼虫、幼体(亚成体)或成体[14]。本研究首先结合形态学与18S 分子标签鉴定了网捕礁栖鱼类的种类,然后采用长棘海星特异性引物检 测了鱼类肠道内容物中的 CoTS-mtCOⅠ 基因片段,以此来鉴别中国南海西沙岛礁中的长棘海星鱼类捕食者。在珠蝴蝶鱼、三间火箭、弓月蝴蝶鱼、红裸颊鲷、东方尖唇鱼和蜂巢石 斑鱼等 6 种鱼的肠道内容物中均检测到 CoTS-mtCOⅠ 基因片段,表明它们具有长棘海星捕食能力。珠蝴蝶鱼、三间火箭和弓月蝴蝶鱼同隶属于蝴蝶鱼科,红裸颊鲷隶属于裸颊鲷科,东方尖唇鱼隶属于隆头鱼科,蜂巢石斑鱼隶属于石斑鱼科。
已有研究表明,蝴蝶鱼科鱼类能摄食长棘海星成体、配子或幼虫[24, 27-28] ,如丝蝴蝶鱼 (Chaetodon auriga) 、胡麻斑蝴蝶鱼(C. citrinellus)和斜纹蝴蝶鱼(C. vagabundus)喜捕食受伤和濒死的长棘海星成体[13, 24, 27-31]。在本研究中,西沙礁区珠蝴蝶鱼和弓月蝴蝶鱼肠道内容物DNA中CoTS-mtCOⅠ 基因片段的检出率为100%,三间火箭肠道内容物 DNA 中检出率均 66% 以上 。前两者的摄食能力明显高于已报道的叉纹蝴蝶鱼(C. auripes) ,这表明珠蝴蝶鱼和弓月蝴蝶鱼可能具有长棘海星摄食偏好性。三间火箭的摄食能力低于珠蝴蝶鱼和弓月蝴蝶鱼,这种差异可能源自它们的食性差异。珠蝴蝶鱼是杂食性鱼类,弓月蝴蝶鱼是珊瑚虫食性鱼类,三间火箭是利用延长的管状吻在珊瑚礁区进行捕食的肉食性鱼类[32-34] 。由此可见,尽管蝴蝶鱼科鱼类能摄食不同生长阶段的长棘海星,但其摄食方式和能力存在明显差异。
本研究在西沙礁区红裸颊鲷、东方尖唇鱼和蜂巢石斑鱼肠道内容物 DNA中的 CoTS-mtCOⅠ 基因片段检出率均≥50%。红裸颊鲷、东方尖唇鱼和蜂巢石斑鱼三者都是典型的礁区肉食性鱼类,同科的鱼类均已发现具有长棘海星摄食能力 。如太平洋裸颊鲷(Lethrinus atkinsoni)、长吻裸颊鲷(L. miniatus)、星斑裸颊鲷(L. nebulosus)三种裸颊鲷科鱼类对长棘海星幼体和 成 体 的 摄 食[25, 29, 35]; 横 带 唇 鱼( Cheilinus fasciatus) 、 波纹唇鱼(C. undulatus)和黑尾海猪鱼(Halichoeres melanurus)等隆头鱼科鱼类对长棘海星成体的摄食[36-38];鞍带石斑鱼对长棘海星幼体的摄食[26] 。这些研究多是通过室内模拟观察到,且未评估鱼类的长棘海星摄食能力。
本研究首次在中国南海西沙礁区发现 6 种长棘海星潜在鱼类捕食者,包括珠蝴蝶鱼、三间火箭、弓月蝴蝶鱼、红裸颊鲷、东方尖唇鱼和蜂巢石斑鱼。这 6 种鱼类的平均摄食水≥50%,其中珠蝴蝶鱼和弓月蝴蝶鱼可能对长棘海星具有摄食偏好性。后续研究应着眼于解析捕食者的捕食效率及其对长棘海星种群的调控机制,为制定预防中国南海长棘海星暴发的策略提供重要科学指导。
Potential fish predator analysis of crown-of-thorns starfish Acanthaster planci in the Xisha reef area using intestinal content DNA
DOI: 10.15886/j.cnki.rdswxb.20250101
- Received Date: 2025-08-02
- Rev Recd Date: 2025-10-04
- Available Online: 2025-10-22
- Publish Date: 2026-01-25
-
Key words:
- Acanthaster cf. solaris /
- predator /
- intestinal content /
- molecular markers /
- mitochondrial gene
Abstract: To understand the fish predators of crown-of-thorns starfish Acanthaster cf. solaris in the Xisha Islands, this study utilized 18S rRNA sequencing and morphological characteristics to identify the fish species collected in April 2023, and then detected the crown-of-thorns starfish mitochondrial cytochrome-C-oxidase subunit Ⅰ (CoTS-mtCOⅠ) gene fragments in fish intestinal content DNA. Analysis of 330 fish revealed 42 species across 37 genera, 24 families and 9 orders. The CoTS-mtCOⅠ fragment was detectable in the intestinal content DNA of six fish species, including Chaetodon kleinii, Chelmon rostratus, Chaetodon lunulatus, Lethrinus haematopterus, Oxycheilinus orientalis, and Epinephelus merra. The detection rate of CoTS-mtCOⅠ gene fragment was 100% in Chaetodon kleinii, while it was only detected in partial samples of the other five species. The six species were firstly reported fish predators of the crown-of-thorns starfish. Phylogenetic analysis based on 18S sequences grouped Chaetodon kleinii, Chelmon rostratus, and Chaetodon lunulatus within the family Chaetodontidae, and placed L. haematopterus, O. orientalis, and E. merra within the order Perciformes. This study contributes to understanding the fish predators of the crown-of-thorns starfish in the coral reef ecosystem of the Xisha Islands, providing a theoretical basis and scientific guidance for the prevention and control of the outbreak of the crown-of-thorns starfish in the South China Sea.
| Citation: | Xing Jiajie, Li Zhonglu, Yu Qiuyu, He Chunlong, Zhou Zhi, Liu Zhaoqun. Potential fish predator analysis of crown-of-thorns starfish Acanthaster planci in the Xisha reef area using intestinal content DNA[J]. Journal of Tropical Biology, 2026, 17(1): 155-164. doi: 10.15886/j.cnki.rdswxb.20250101

|
 DownLoad:
DownLoad: