
-
槟榔(Areca catechu L.)属于棕榈科槟榔属,为常绿乔木,位居中国四大南药之首,是海南省第一大特色经济作物,在海南种植面积接近20万hm2[1-2]。近年来,黄化病严重威胁槟榔种植业,已经成为限制海南地区槟榔产业发展最重要的因素[3],其中,槟榔黄化相关病毒(areca palm velarivirus 1,APV1)被确定为关键病原之一[4-6]。APV1典型症状包括叶片黄化、植株矮化、果实畸形等,最终导致产量锐减甚至绝收[4]。
APV1属于长线形病毒科(Closteroviridae)隐症病毒属(Velarivirus)[7],是单分体正义单链RNA(plus sense single-stranded RNA, +ssRNA)植物病毒,其病毒粒子呈弯曲的丝状,长度为2 000~3 500 nm[4,8-9]。APV1基因组具有高度保守性,其11个开放阅读框(open reading Frame, ORF)呈现功能模块化分布特征[10]。5’端ORF1a/b编码病毒复制必需的多聚酶复合体,ORF2编码4.3 kDa疏水多肽p4,ORF3编码具有分子伴侣功能的Hsp70同源蛋白。结构蛋白编码区中,ORF5编码60 kDa的p60组装调控因子,ORF6/ORF7分别编码主要衣壳蛋白(coat protein,CP)与次要衣壳蛋白(minor capsid protein,CPm)。其中,CP特异性识别并包裹病毒基因组RNA(gRNA)3’端约95%区域,与CPm协同完成核衣壳的层级组装,而CPm则负责gRNA尾部5%区域的包裹,是病毒能否侵染宿主最重要的蛋白,并且发现CP蛋白分别与sHSP和DnaJB13互作[11];除了以上编码蛋白,ORF4、ORF8、ORF9、ORF10未能比对到相似的蛋白,其功能未知。在植物与病毒长期的协同进化“军备竞赛”进程中,几乎全部植物病毒均可编码RNA沉默抑制子(viral suppressors of RNA silencing,VSR)。该类蛋白的主要作用是抵御宿主的防御反应,从而助力病毒完成侵染、复制及系统性扩散的生命过程[12]。得益于长线形病毒复杂的基因组提供的负载能力,部分成员能够编码多个VSR,且其通常定位在基因组上3’端可变区域。长线形病毒编码的多个VSR在功能上表现出分工和协作,例如柑橘衰退病毒(citrus tristeza virus,CTV)编码3个VSR,其中p20在本氏烟上抑制局部和系统沉默,而CP仅靶向系统沉默,p23仅靶向局部沉默[13];p20和p23的共同表达增强了对局部沉默的抑制[14]。菠萝凋萎病毒2(pineapple mealybug wilt associated virus-2,PMWaV-2)编码4个VSR,p20和p22都靶向系统沉默[15]。尽管APV1的基因组序列已被解析[16],但其3’端的ORF4、ORF8、ORF9、ORF10编码的p21、p26、p18、p25蛋白功能仍然未知。p26蛋白大小为25.7 kDa,海南大学黄惜教授团队前期研究表明p26是RNA沉默抑制子,也是重要的致病因子(未发表)。因此,揭示p26蛋白的互作蛋白对解析APV1的致病机制具有重要意义。
酵母双杂交(yeast two-hybrid,Y2H)技术是检测蛋白质间相互作用(protein-protein interaction,PPI)的经典方法[17]。其原理基于利用转录因子的模块化特性:将报告基因(如HIS3、LacZ)置于该转录因子(如Gal4)结合位点(UAS)下游;同时,将转录因子的DNA结合域(DNA-binding domain,BD)与诱饵蛋白(Bait)融合,激活域(activation domain,AD)与潜在的互作蛋白(猎物,Prey)融合。只有当诱饵蛋白与猎物蛋白发生特异性相互作用时,才能将BD和AD在空间上拉近,重构出有功能的转录因子,从而激活下游报告基因的表达,使酵母在特定筛选培养基上生长或显色。该技术因其直接在活细胞(酵母)内检测互作、灵敏度高、可高通量筛选文库等优势,被广泛应用于构建病毒−宿主互作研究。例如,利用水稻条纹病毒(RSV)的VSR p3蛋白作为诱饵筛选烟草cDNA文库,鉴定出宿主互作蛋白NbP3IP[18]。
基于上述研究背景,本研究选取APV1的p26蛋白作为诱饵,借助酵母双杂交技术对槟榔cDNA文库进行筛选,并鉴定与p26存在互作关系的宿主蛋白。后续将针对所筛选获得的宿主因子开展生物学功能解析研究,以期为深入揭示p26蛋白在APV1病毒侵染寄主过程中的作用机制提供理论支撑。
-
本研究采用槟榔黄化病病叶,源自海南省三亚市崖州区(18°21′32.9″ N, 109°10′2.5″ E)。DH5α大肠杆菌感受态细胞、Y2HGold与Y187酵母感受态细胞均采购于上海唯地公司。酵母双杂交系统配套载体pGBKT7(诱饵载体)及pGADT7(猎物载体)购自大连宝生物公司。槟榔cDNA文库由邢增宇等[11]前期构建。所有引物的合成工作委托上海生工生物工程公司完成(详见表1)。
引物名称 Primer 引物序列 Sequence pGBKT7-p26-F CAGAGGAGGACCTGCATATGGAAGATCTTGACACG pGBKT7-p26-R TAGTTATGCGGCCGCTGCAGTCATAATTTTATTTTTC CHCHD2-F GTACCAGATTACGCTCATATGATGCCTCGCCGAAGCTC CHCHD2-R CAGCTCGAGCTCGATGGATCCTTAAGCACCCAGTGTAG AD-F AATACCACTACAATGGATGATG AD-R ATGGTGCACGATGCACAGT BD-F GTGCGACATCATCATCGGAAG BD-R CTAAGAGTCACTTTAAAATTTGTA TACAC cLUC-p26-F CGGGGCGGTACCCGGGATCCAGAGGACTTGGACACGAAGGT cLUC-p26-R CGAAAGCTCTGCAGGTCGACCTACAGCTTGATCTTGCCCA nLUC-AcCHCHD2-F ACGAGCTCGGTACCCGGGATCCATGCCTCGCCGAAGCTCTG nLUC-AcCHCHD2-R GGTGCTACACTGGGTGCTGTCGACCAGATCTCGTACGCGT Table 1. Primer sequence information
-
利用天根生化科技公司的总RNA提取试剂盒(#DP441)对含有APV1病毒的槟榔叶片进行RNA提取(RNA质量利用吸光度法测量,OD260/OD280为1.8~2.0)。使用山东思科捷生物技术有限公司的第一链cDNA合成试剂盒(#AG0304),参考说明书进行cDNA第1链的合成。
-
参考NCBI数据库中APV1-WNY株(GenBank登录号:MK956940.2)的基因序列信息,设计特异性引物pGBKT7-p26-F/R,以上述得到的cDNA为模板进行PCR扩增,纯化回收后得到目的片段。采用南京诺唯赞公司的重组一步克隆试剂盒(#C115),构建诱饵表达载体pGBKT7-p26,PCR验证后经生工生物测序,序列正确的样品于−20 ℃保存。
-
毒性检测:pGBKT7-p26质粒与pGBKT7空载质粒分别转入酵母Y2H Gold,涂布于SD/-Trp固体平板培养基并于2×YPDA液体培养基培养,稀释菌液点板于SD/-Trp固体平板后,30 ℃培养1~2 d观察生长情况。
自激活检测:将pGBKT7-p26和pGADT7空载共转化至Y2H Gold,涂布DDO(SD-Trp/-Leu)固体平板,筛选阳性单克隆,稀释菌液后点板于DDO、QDO/X(SD-Trp/Leu/-His/-Ade+X-α-gal)平板上(本文所用X-α-gal浓度均为20 mg·mL−1),30 ℃培养3~5 d观察生长情况。
-
酵母双杂交筛库实验参考北京宝日医生物技术有限公司的酵母双杂交操作手册[11],并调整具体流程如下:4~5 mL pGBKT7-p26诱饵菌株与1 mL pGADT7-cDNA文库(文库来源于邢增宇等[11])混合,于45 mL 2×YPDA液体培养基30 ℃共培养24 h后涂布于30个TDO/X(SD/-Trp/-Leu/-His+X-α-gal)平板,30 ℃条件下培养3~5 d,待长出约800个克隆后筛选蓝色菌落;阳性菌落依次在TDO/X和QDO/X复筛3次和5次,确认互作特异性,提取酵母质粒转化DH5α,PCR验证阳性菌液后,经生工生物公司测序,并通过NCBI网站(
https://www.ncbi.nlm.nih.gov/ )分析互作基因。 -
将同源物种的CHCHD2碱基序列在槟榔转录组数据库中BLAST,选取同源性最高的基因设计上下游引物CHCHD2-F/R,使用大连宝生物(#R050)高保真酶Prime STARGXL DNA Polymerase扩增目标产物,连接pGADT7载体(pGADT7-AcCHCHD2),转化DH5α,PCR验证后经生工生物测序。序列正确的样品于−20 ℃保存。
-
对槟榔候选基因CHCHD2进行回转验证实验[19],具体步骤如下:将pGBKT7-p26与pGADT7-AcCHCHD2质粒共转化至Y2H Gold中,涂布于DDO(SD/-Trp/-Leu),筛选阳性单克隆的菌液稀释后分别点板于DDO和QDO/X平板,30 ℃培养3~5 d观察菌落生长状况。
-
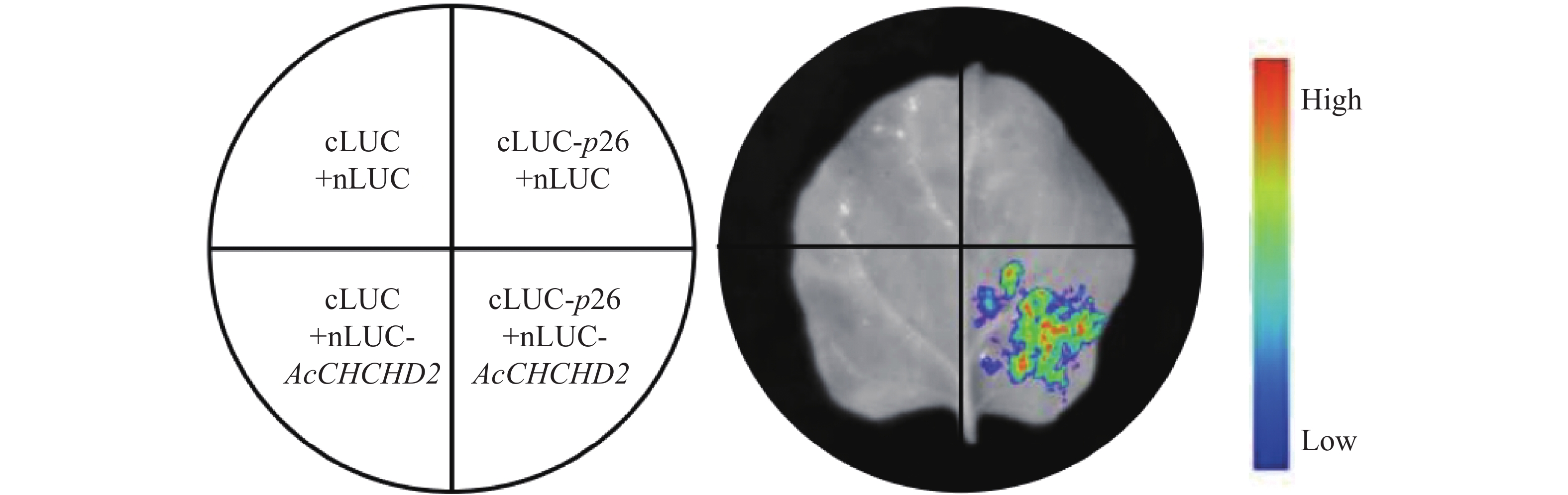
在萤光素酶互补成像(luciferase complementation imaging, LCI)分析中,将p26的编码序列克隆至萤光素酶(luciferase, LUC)的C端,构建cLUC-p26,同时将AcCHCHD2编码序列克隆至LUC的N端,构建nLUC-AcCHCHD2(所用引物见表1)。通过农杆菌浸润法,使重组构建载体在本氏烟草叶片中瞬时表达。在侵染后48 h收获叶片,用1 mmol·L-1 D-萤光素钾盐涂抹于叶片背面避光孵育8~10 min,然后使用NightShade LB 985植物活体成像系统(德国贝托尔德技术公司)进行检测,并拍摄图。
-
蛋白质理化性质通过ExPASy ProtParam(
https://web.expasy.org/protparam/ )分析;亲疏水性利用ProtScale(https://web.expasy.org/protscale/ )预测;同源性比对采用NCBI BLAST,本地比对用Bioedit软件;跨膜结构域通过TMHMM Server v.2.0(https:/services.healthtech.dtu.dk/service.php?TMHMM-2.0 )分析;信号肽预测使用SignalP 4.1 Server(https://services.healthtech.dtu.dk/services/SignalP-4.1/ );二级结构通过SOMPA网站(https://npsa.lyon.inserm.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html )预测;亚细胞定位利用PSORT网站(https://wolfpsort.hgc.jp/ )分析。 -
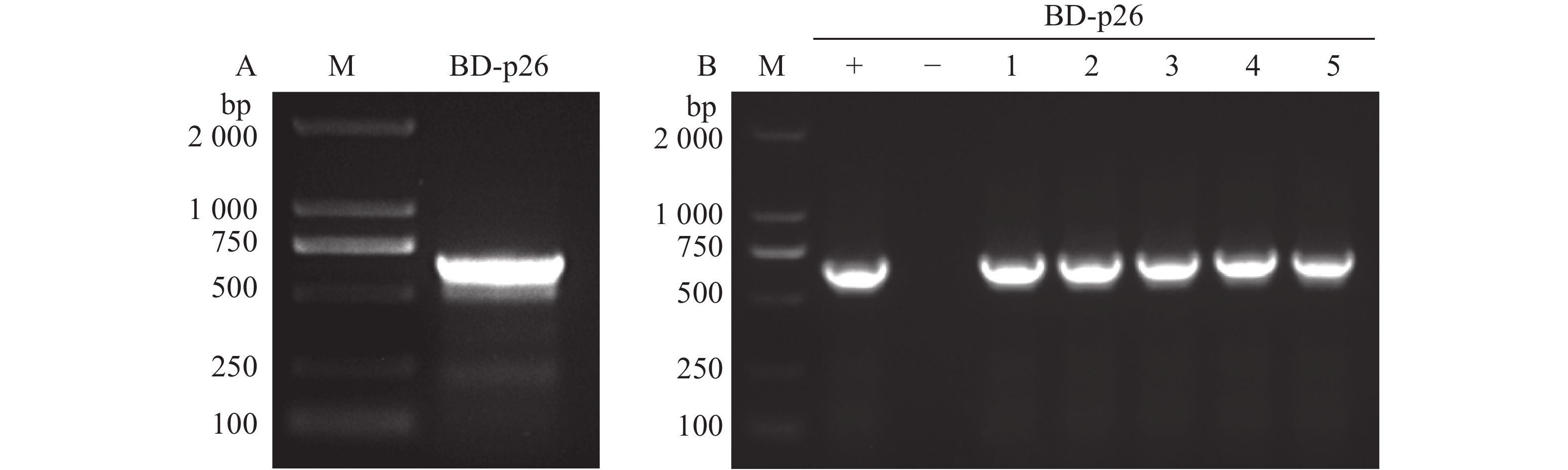
以槟榔黄化病病叶cDNA为模板,用特异性引物pGBKT7-p26-F/R经高保真酶PCR扩增APV1-p26,产物电泳检测条带正确(图1-A);将目的片段连接pGBKT7载体后转化DH5α,挑取5个单克隆菌落PCR检测,电泳结果与阳性对照一致(图1-B),表明成功构建pGBKT7-p26诱饵载体。
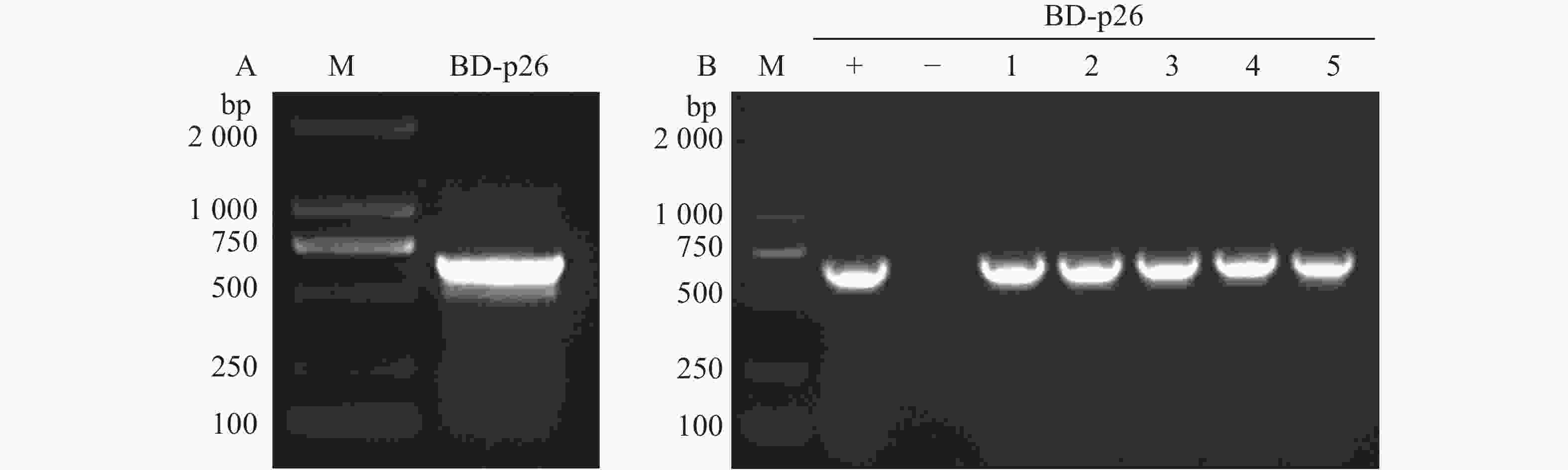
Figure 1. Construction of the pGBKT7-p26 bait expression vector
-
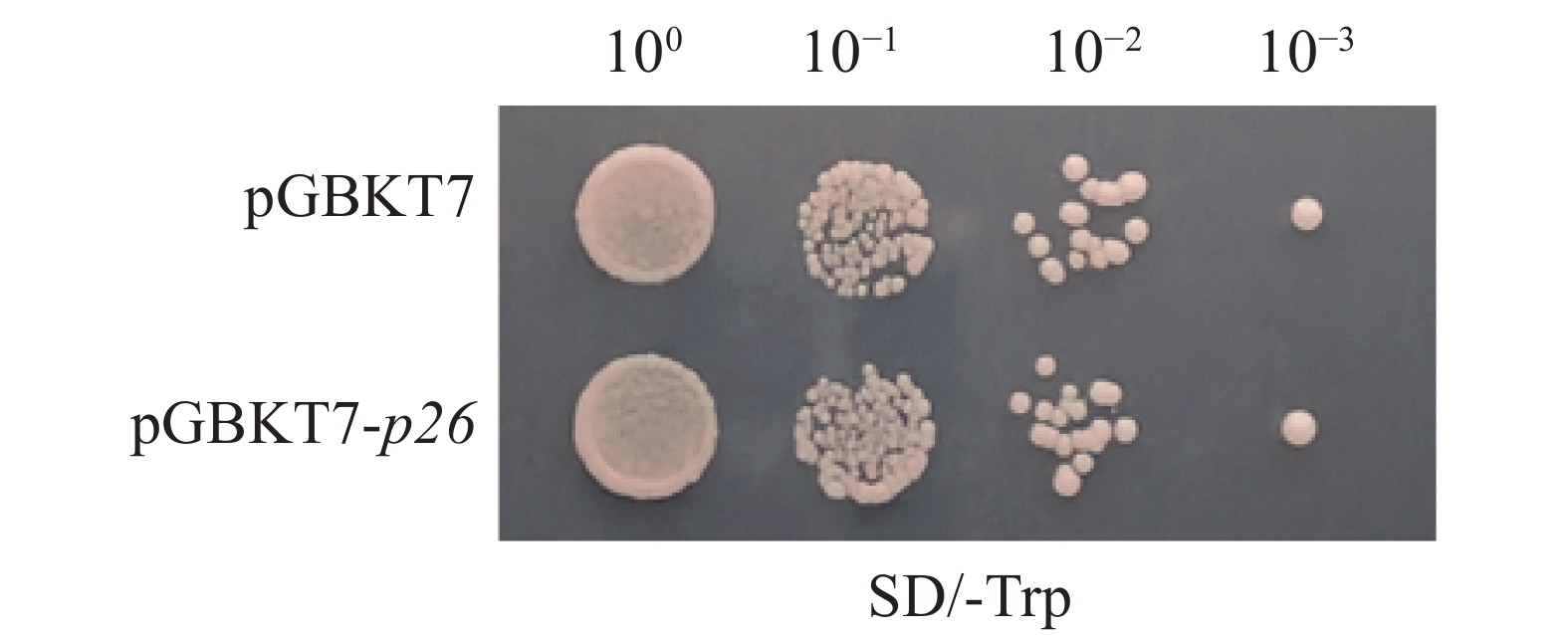
将pGBKT7-p26重组质粒及pGBKT7空载体分别转化Y2H Gold感受态细胞,结果显示两者单克隆数量及直径无显著差异,证实pGBKT7-p26诱饵载体对酵母细胞无毒性(图2)。
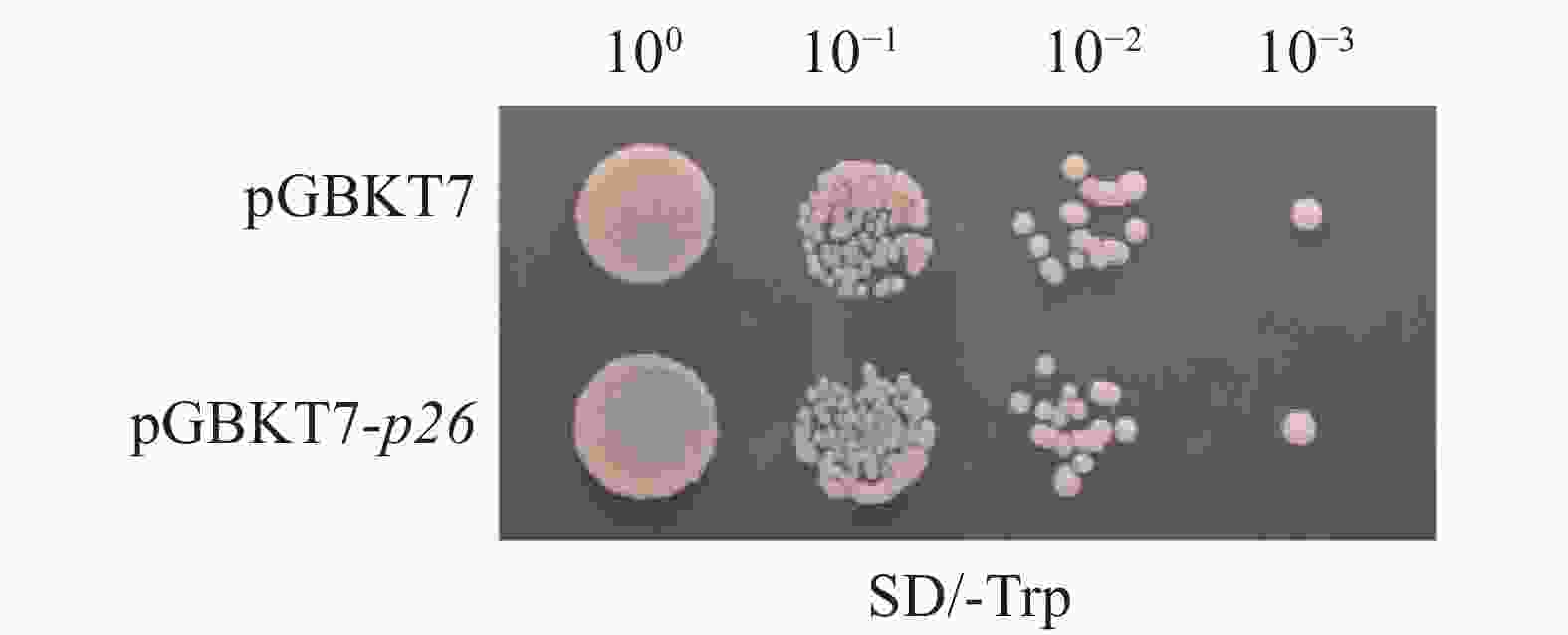
Figure 2. Toxicity detection of pGBKT7-p26
-
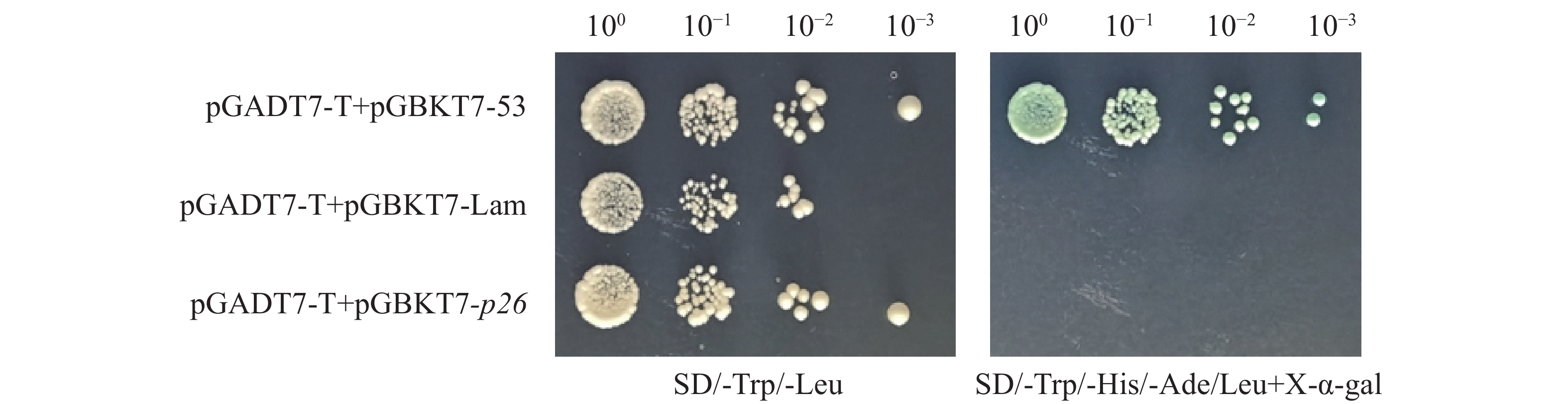
将pGBKT7-p26与pGADT7空载体共转化Y2H Gold,营养缺陷型筛选结果显示:实验组(pGBKT7-p26 + pGADT7-Empty)与阴性对照(pGBKT7-Lam + pGADT7-T)在DDO平板生长但QDO/X平板不生长;阳性对照(pGBKT7-53 + pGADT7-T)在DDO平板生长且QDO/X平板显蓝。这表明pGBKT7-p26无自激活活性,可用于酵母双杂交筛库(图3)。

Figure 3. Self-activation detection of pGBKT7-p26
-
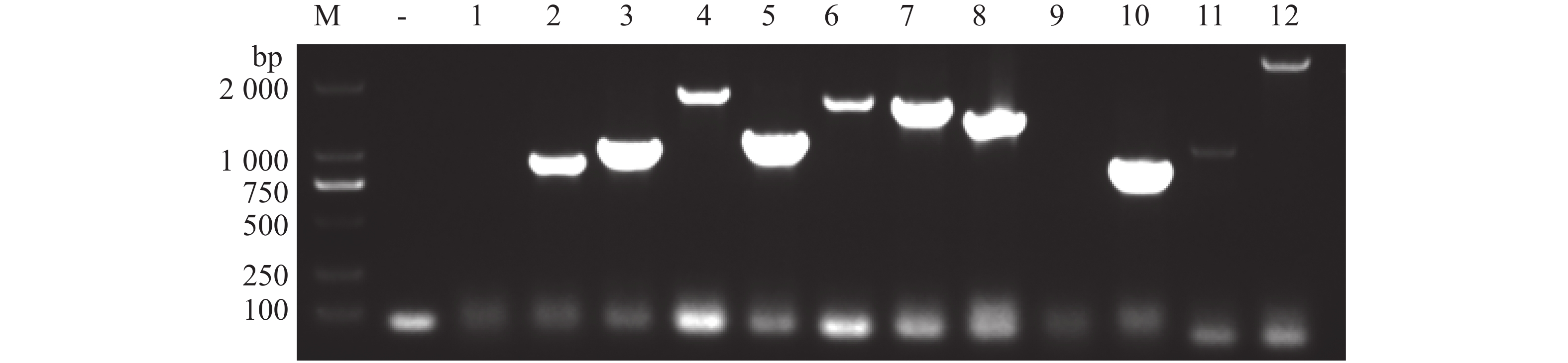
将含pGBKT7-p26的Y2HGold菌株培养至对数期(OD600≈0.8),收集菌体重悬于2×YPDA,与pGADT7-cDNA文库共孵育20 h后经0.5×YPDA洗涤,涂布TDO/X平板。挑取蓝色菌落依次经TDO/X(3轮)和QDO/X(5轮)复筛(图4)。从QDO/X平板取12个稳定显色的单克隆,提酵母质粒转化DH5α,菌液PCR获10个阳性克隆(图5)。测序结果经NCBI BLAST及本地槟榔转录组比对,筛选出6个高可信度候选互作蛋白(表2)。

Figure 4. Screening of host proteins interacting with pGBKT7-p26
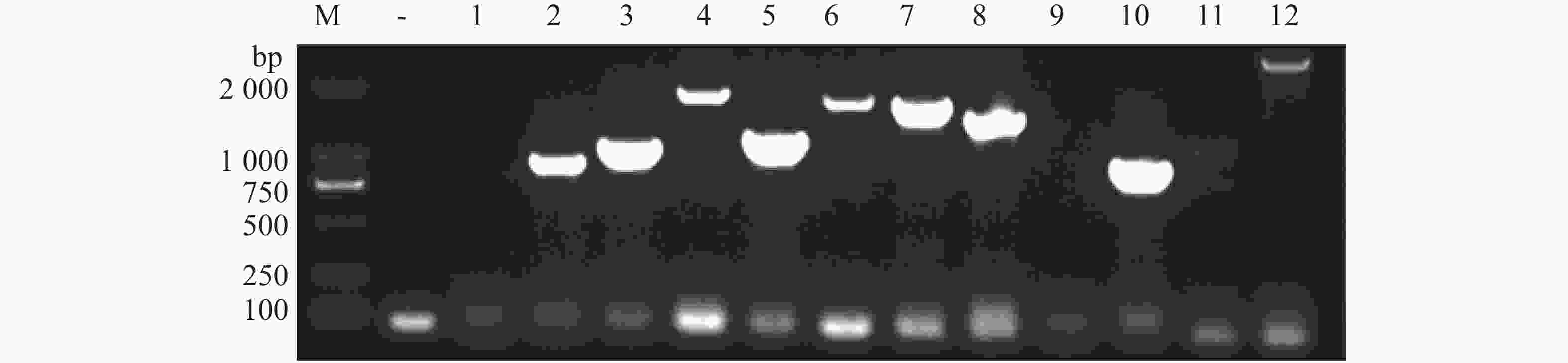
Figure 5. Results of Escherichia coli bacterial liquid PCR detection (Verification of positive clones by blue-white screening)
编号No. Genebank登录号 GenBank accession number 基因注释 Gene annotation 1 XM_010924085.3 油棕Ras相关蛋白 RABE1c
Elaeis guineensis ras-related protein RABE1c2 XR_ 605948.4 海枣未表征蛋白
Phoenix dactylifera uncharacterized protein3 XM_010924987.3 油棕未表征蛋白 C6C3.02c
Elaeis guineensis uncharacterized protein C6C3.02c4 XM_010910619.3 海枣未表征蛋白
Elaeis guineensis uncharacterized protein5 XM_010933347.2 油棕应激蛋白 PHOS34
Elaeis guineensis universal stress protein PHOS346 XM_010924311.3 油棕叶绿体铁蛋白- 4
Elaeis guineensis ferritin-4, chloroplasticTable 2. Information on candidate interacting proteins of p26
-
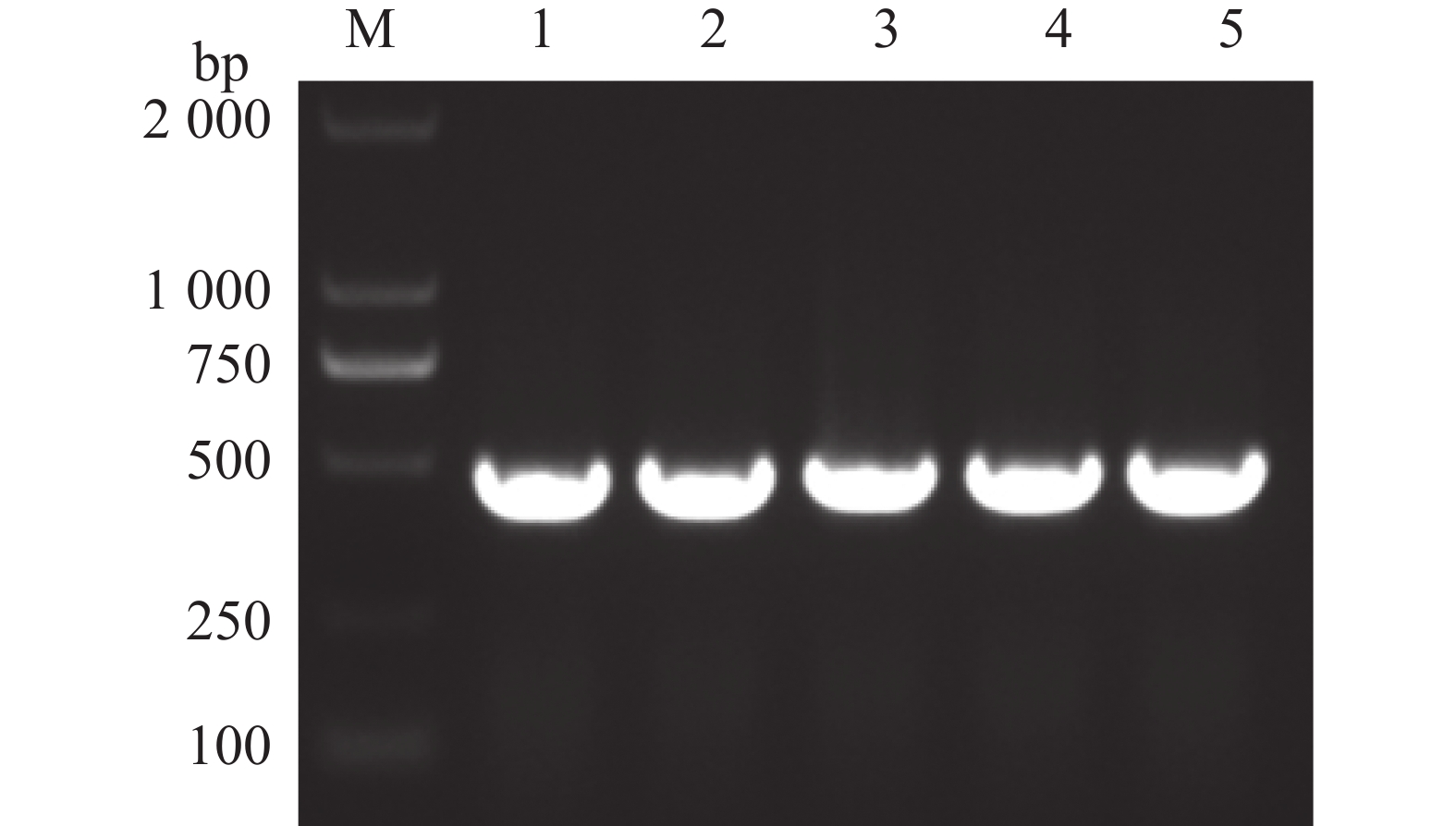
为了进一步验证候选互作蛋白与p26蛋白互作的准确性,选取表2中槟榔候选3号未知功能蛋白进行酵母双杂交回转验证。通过结构域预测分析,发现该基因编码产物具有典型的卷曲螺旋结构域(coiled-coil-helix-coiled-coil-helix domain),与动物卷曲螺旋结构域蛋白2(CHCHD2)家族成员具有显著序列相似性,故命名为槟榔卷曲螺旋结构域蛋白2(AcCHCHD2,coiled-coil-helix-coiled-coil-helix domain containing 2)。该序列在槟榔转录组数据库中进行BLAST比对,获取同源性最高的编码序列。设计特异性引物CHCHD2-F/R(表1),以槟榔cDNA为模板扩增AcCHCHD2基因,构建pGADT7-AcCHCHD2重组载体。阳性克隆经测序验证(图6)后,与pGBKT7-p26共转化Y2H Gold。转化子在DDO培养基上均生长,表明诱饵和猎物载体已整合并表达。在QDO/X选择性培养基上,除阴性对照外的所有实验组均生长良好且显色,证实p26与AcCHCHD2在酵母细胞中存在特异性相互作用(图7)。
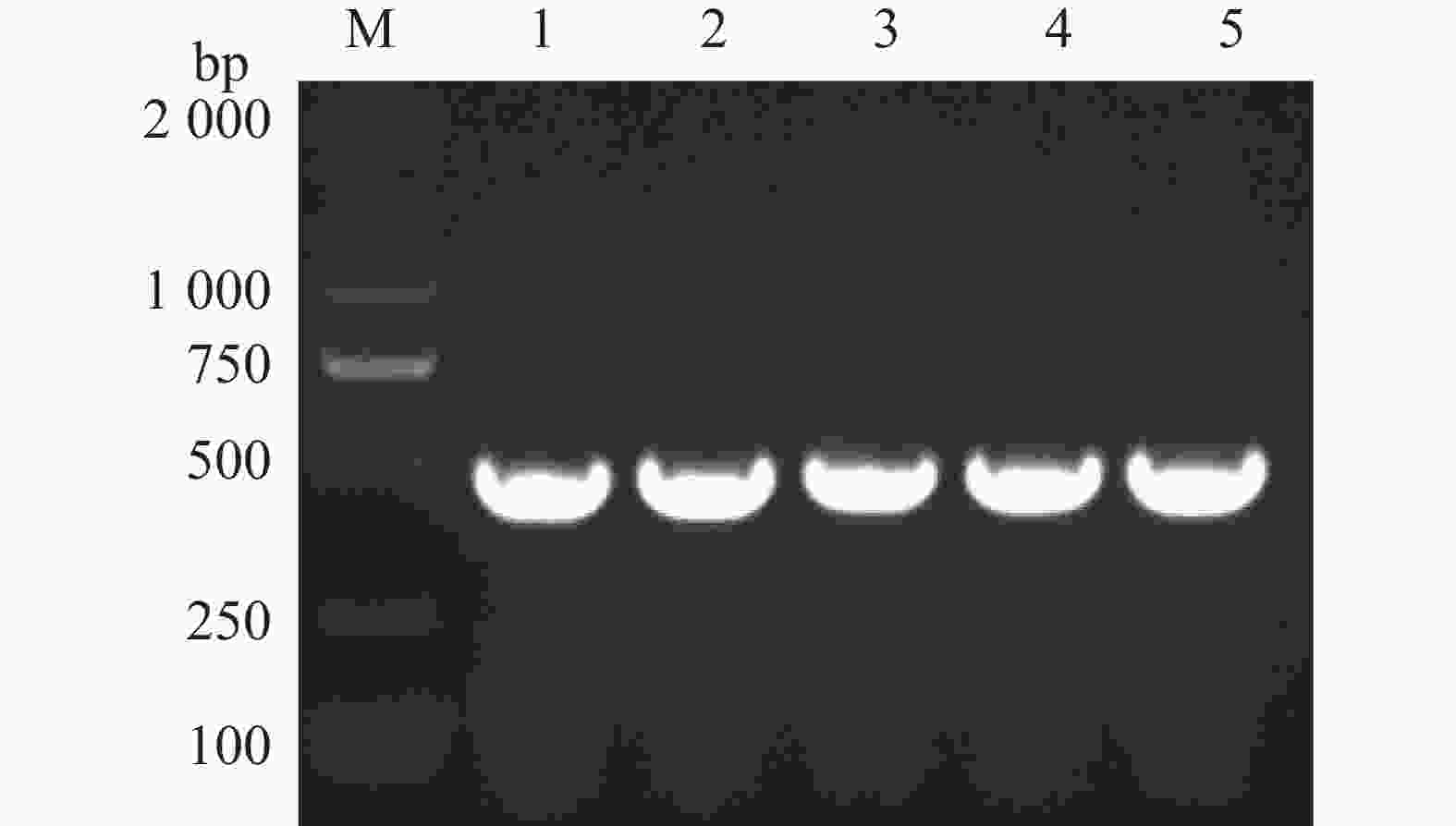
Figure 6. PCR verification of areca palm CHCHD2 protein gene cloned from Escherichia coli

Figure 7. Rotation validation of p26 protein versus CHCHD2 protein
-
为了验证酵母体系中的试验结果,将互作蛋白p26、CHCHD2基因全长序列分别构建到cLUC、nLUC载体后,转入农杆菌GV3101感受态细胞,之后注射到本氏烟叶片,通过生物活体成像仪检测发现p26蛋白和CHCHD2蛋白在植物体内存在明显的相互作用(图8),进一步验证了p26蛋白和CHCHD2蛋白的互作关系。
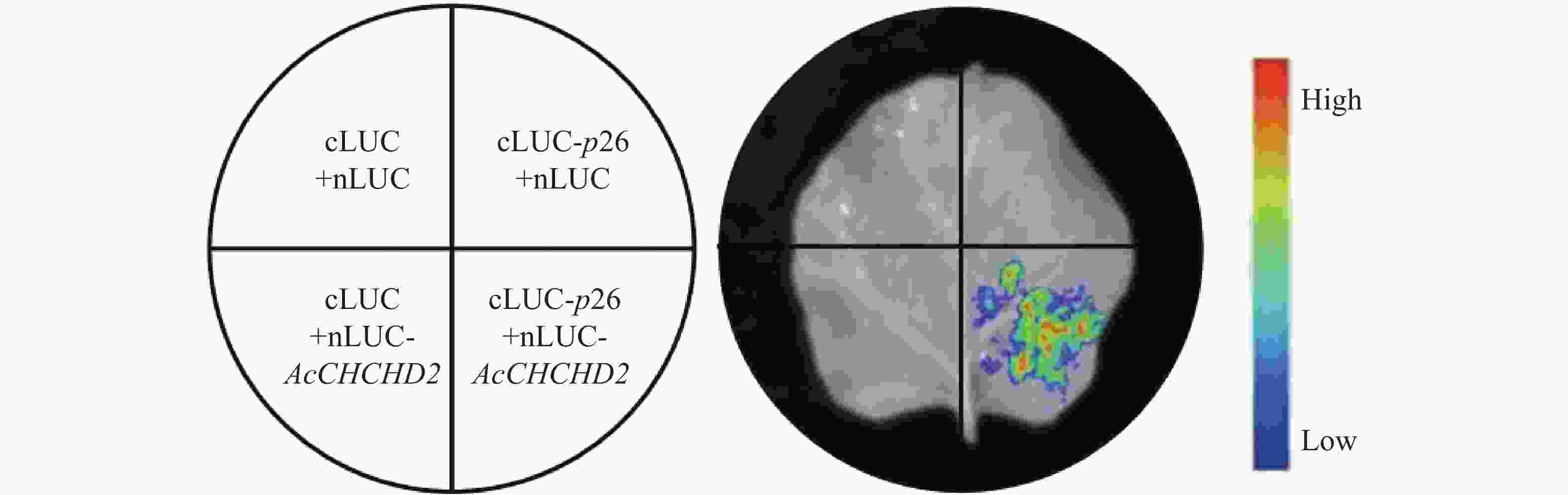
Figure 8. Interaction between p26 and CHCHD2 in LCI experiment
-
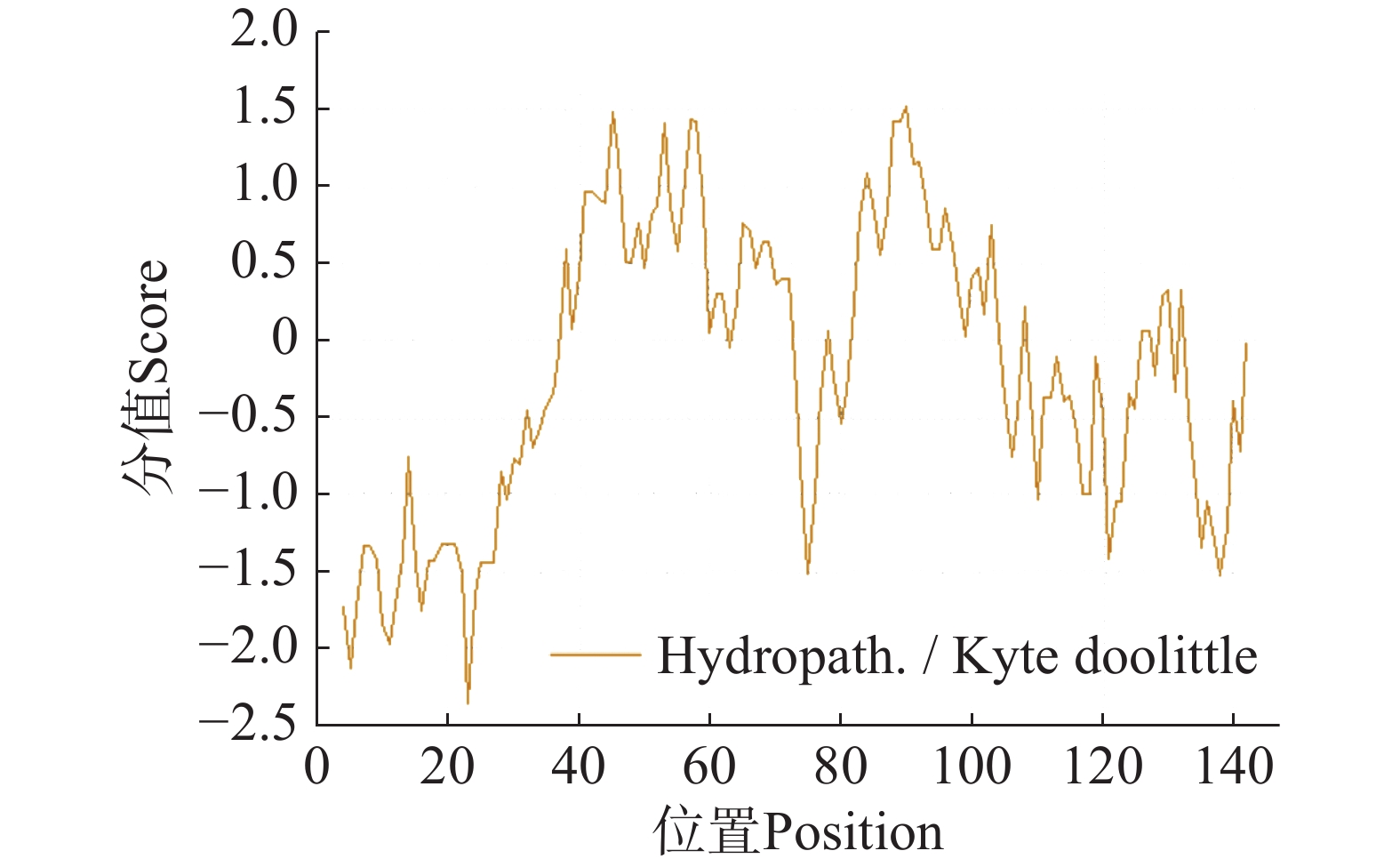
采用ExPASy生物信息学平台对CHCHD2蛋白理化特性进行系统解析。分析显示,该蛋白化学组成为C628H1 007N201O198S8,编码序列长度441 bp,分子质量为14.8 kDa,原子总数达2 042个,理论等电点(pI)为9.24。氨基酸组成分析表明其富含丙氨酸(Ala, 16.3%)与甘氨酸(Gly, 12.2%),电荷分布呈现净正电荷特性(正电荷残基12个,负电荷残基8个)。理化参数显示脂溶指数62.59提示适度亲脂性,不稳定性指数65.31(阈值<40)明确归类为不稳定蛋白(图9)。
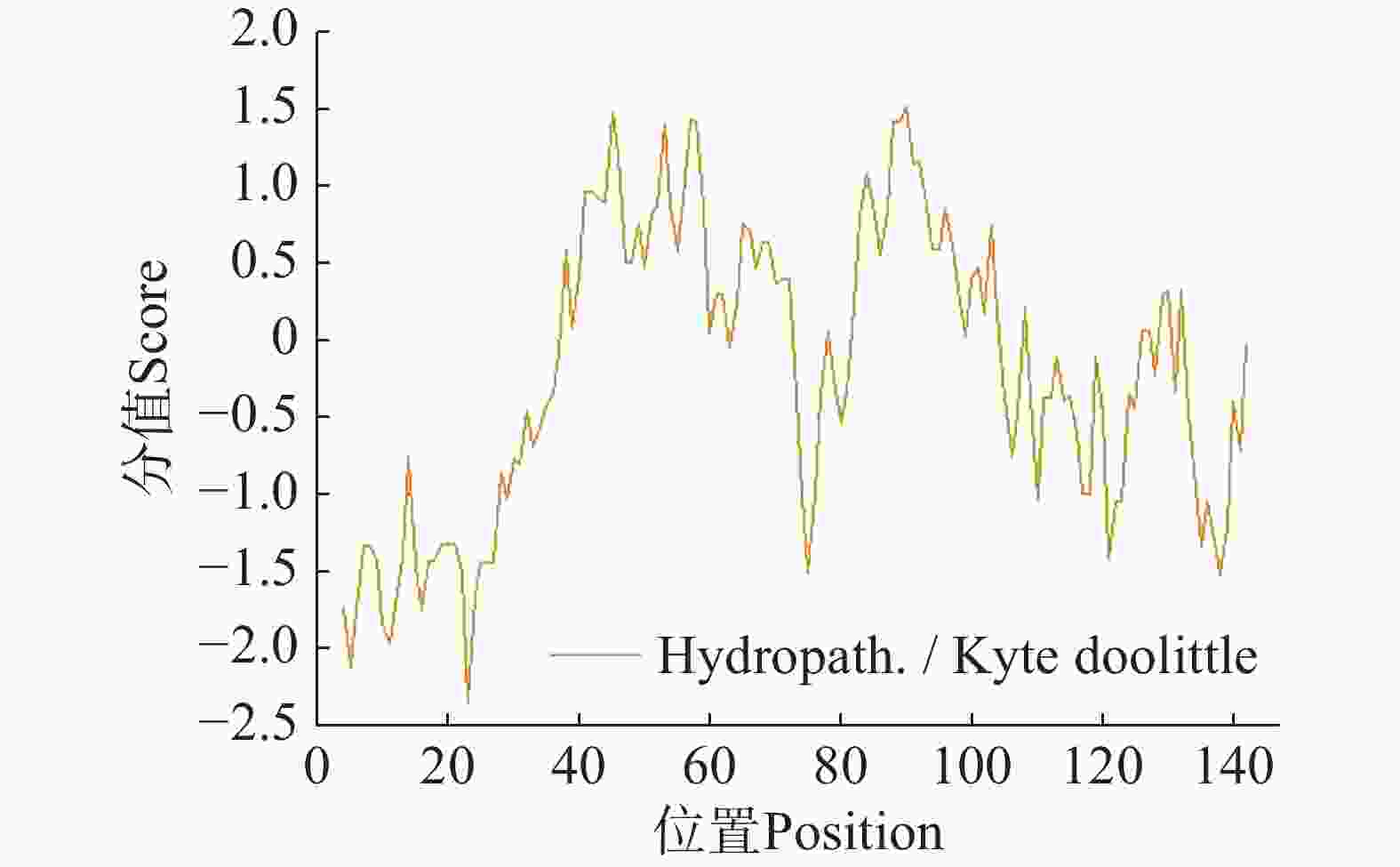
Figure 9. Hydrophilic/hydrophobic prediction of candidate protein CHCHD2
ProtScale亲疏水性分析显示,丙氨酸(Ala)疏水性最强(1.522),脯氨酸(Pro)亲水性最强(−2.356)。整体亲水性残基比例显著高于疏水性残基(图9),表明CHCHD2为亲水性蛋白。
-
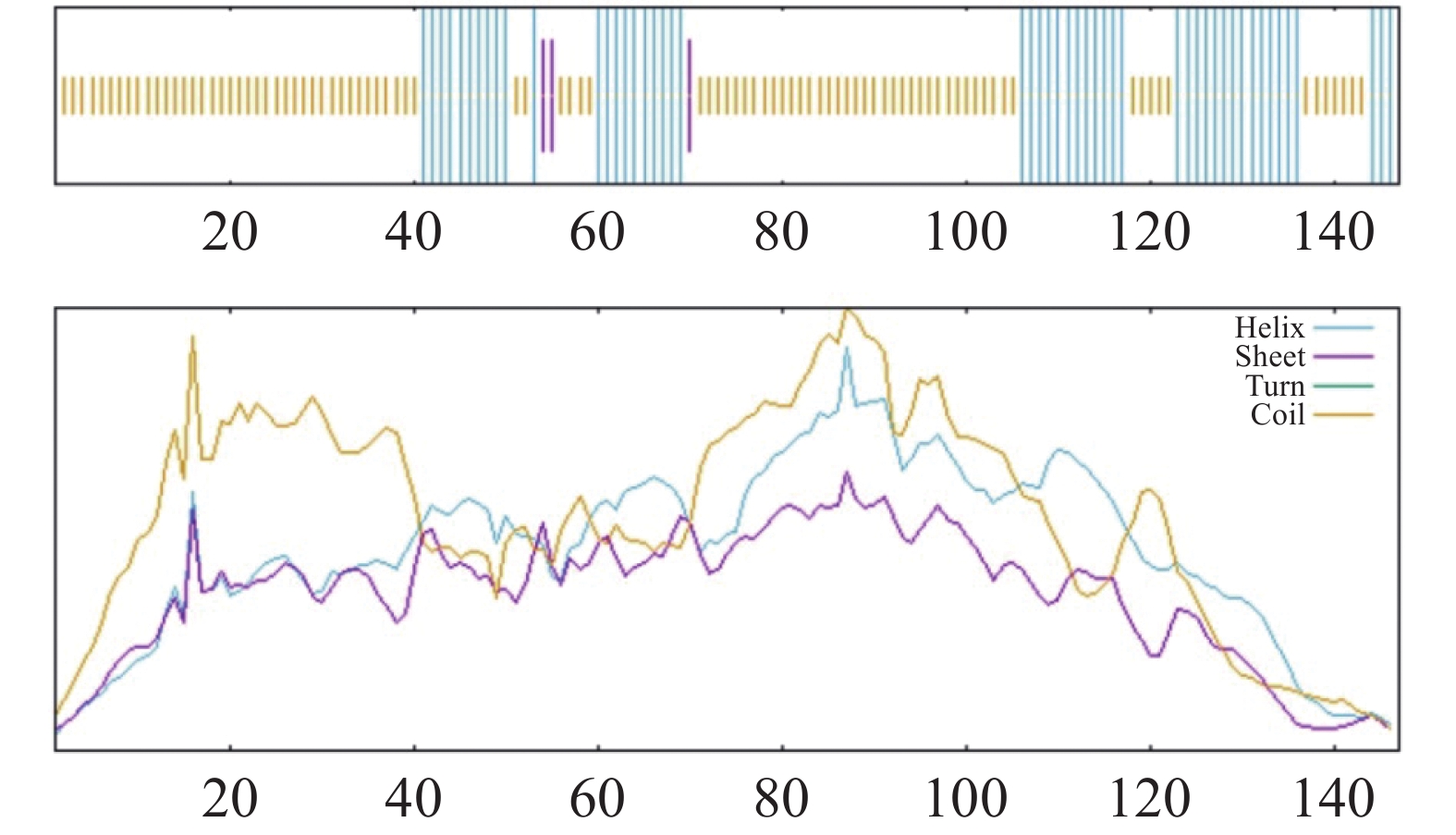
利用SOPMA工具预测CHCHD2的二级结构,显示无规则卷曲(random coil)占比63.95%,α-螺旋(alpha helix)占34.01%,β-折叠(Beta-sheet)占2.04%(图10)
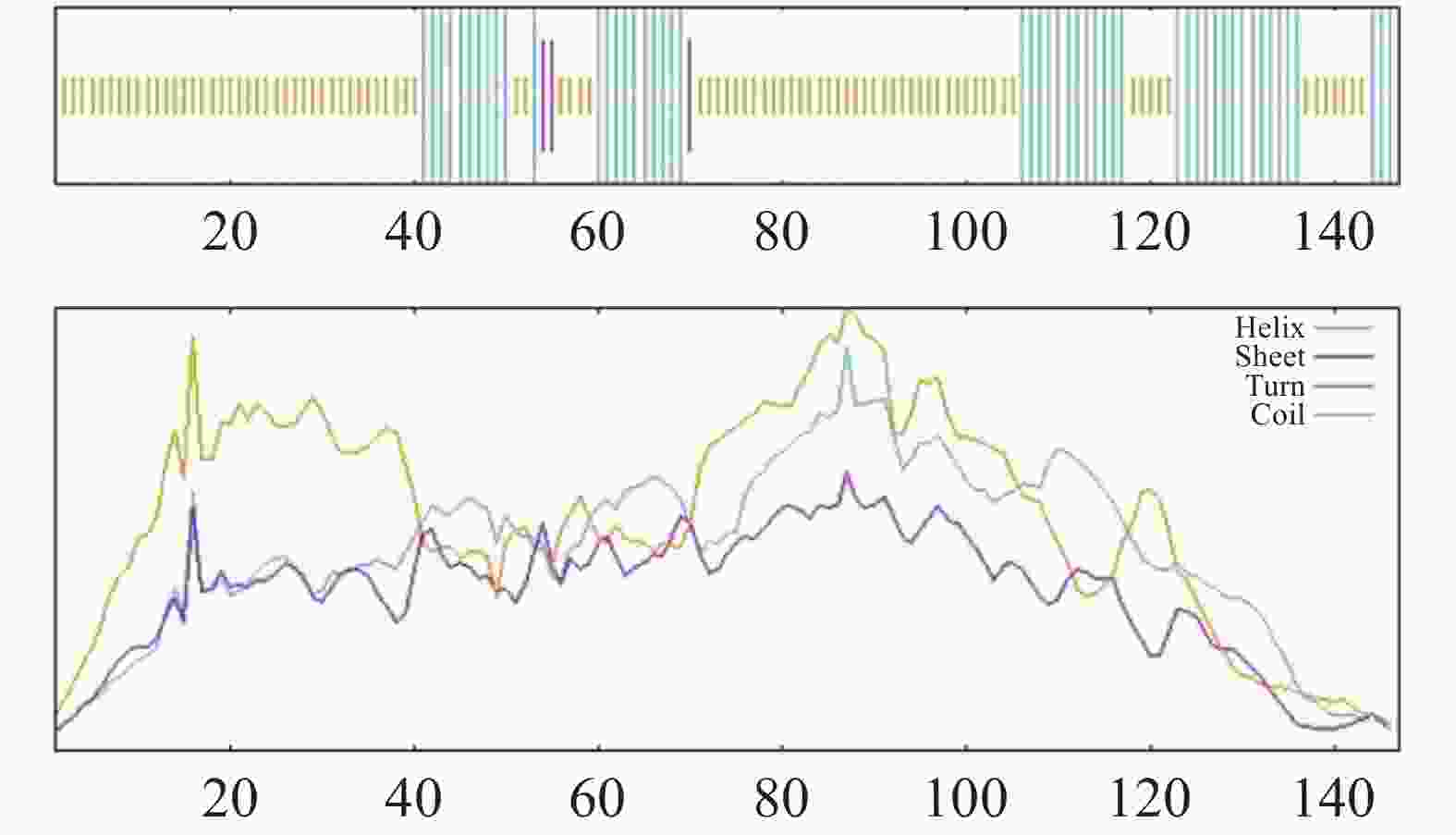
Figure 10. Secondary structure analysis of candidate protein CHCHD2
-
通过SignalP 4.1 Server、PSORT和TMHMM在线工具分别对CHCHD2进行信号肽、亚细胞定位和跨膜结构域分析。结果显示,该蛋白不存在信号肽序列,亚细胞定位预测表明其主要定位于叶绿体,且不含跨膜结构域。
-
APV1属于长线形病毒科,其基因组包含11个开放阅读框(ORF),其中ORF8编码的p26蛋白功能尚未明确。值得注意的是,长线形病毒科的多个成员编码的VSR通常位于基因组3’端可变区[18],而APV1的p26蛋白定位于此区域,提示其可能具有VSR功能,也是APV1的关键致病因子[4]。然而当前对于p26蛋白参与APV1侵染过程,以及植物如何抵御APV1侵染的机制尚不清楚。在植物病毒研究中,Y2H技术揭示了众多病毒与宿主蛋白关键的互作关系,有助于对病毒侵染研究。例如,以黄瓜花叶病毒(CMV)2b蛋白为诱饵筛选拟南芥文库,则发现其与Argonaute 4(AGO4)直接互作[20]。基于Y2H的研究系统性地揭示了多种植物病毒蛋白与宿主因子的互作关系,为深入理解病毒致病机理和挖掘抗病基因提供了重要的理论依据。因此本研究利用Y2H技术,以APV1的p26蛋白为诱饵,筛选得到6个与p26蛋白互作的寄主因子。
在候选蛋白中已知功能蛋白仅有3个为Ras相关蛋白(RABE1c),叶绿体铁蛋白(ferritin-4)和应激蛋白(PHOS34)。RABE1c作为ABA信号正调控因子,通过增强保卫细胞离子转运的ABA敏感性,缩小气孔开度并提高抗旱性[21]。病毒蛋白p26可能通过靶向RABE1c(膜运输)和ferritin-4(铁代谢)促进侵染[22],类似机制见于番茄丛矮病毒p19结合RAB5调控囊泡运输[23],及水稻条纹病毒NS3抑制铁蛋白引发氧化损伤[24]。因此APV1可能采用类似的“劫持宿主通路”策略,需要后续验证是否存在互作。
此外,本研究还筛选到3个未知功能蛋白,其中3号未知功能蛋白经序列比对和结构域分析,被鉴定为卷曲螺旋结构域蛋白CHCHD2。CHCHD2是一类保守的线粒体/叶绿体蛋白,属于卷曲螺旋功能结构域蛋白质家族,在动物中参与氧化应激调控[25],与包括帕金森病在内的多种神经退行性疾病相关,对细胞的正常功能维持起重要作用,多数在线粒体中参与调节线粒体代谢并调节呼吸链成分表达,而在应激情况下其转位至细胞核中调控自身以及细胞色素氧化酶等基因的转录[26],而在植物中的功能尚未明确。本研究首次发现其与病毒蛋白p26互作,暗示APV1侵染过程中可能影响CHCHD2的功能,从而影响槟榔的叶绿体。回转验证实验和LCI实验(图7)进一步证实p26与CHCHD2的特异性互作,排除了筛库过程中的随机结合,为后续体内外互作验证(如Co-IP)提供了坚实的前期证据。
本研究的生物信息学预测显示,槟榔CHCHD2蛋白富含丙氨酸(16.3%)和甘氨酸(12.2%),具有强亲水性(图7),且二级结构以无规则卷曲(63.95%)和α-螺旋(34.01%)为主(图8),这类柔性结构常作为蛋白互作的核心区域,提示其可能通过柔性结构域与p26结合。此外,亚细胞定位预测表明CHCHD2主要定位于叶绿体(无信号肽和跨膜结构域)。据报道,辣椒轻斑驳病毒(pepper mild mottle virus,PMMoV)侵染辣椒可诱导叶绿体外膜蛋白OMP24上调。OMP24过表达触发核周叶绿体聚集及活性氧(ROS)积累,激活叶绿体向细胞核逆行信号(CRS)通路,增强宿主基础免疫以抵御病毒。作为反防御策略,PMMoV外壳蛋白(CP)与OMP24互作,干扰其自身互作,从而抑制OMP24介导的抗病毒免疫[27]。此外,中国番茄黄化曲叶病毒(tomato yellow leaf curl China virus,TYLCCNV)病因子βC1靶向叶绿体蛋白OSB1。βC1-OSB1互作导致OSB1被错误定位至细胞质,阻碍其进入叶绿体并破坏其与叶绿体定位蛋白ALD1的结合,进而促进ALD降解,阻断免疫信号分子哌啶甲酸(PIP)的合成,最终抑制植物免疫[28]。因此推测,APV1 p26可能通过靶向CHCHD2干扰叶绿体功能,破坏宿主ROS稳态,从而削弱抗病毒防御。CHCHD2作为新发现的APV1互作因子,其功能解析可为植物抗病毒育种提供新靶点。
本研究通过酵母双杂交技术筛选到了APV1 p26蛋白的6个候选互作宿主蛋白,并鉴定了CHCHD2为p26的互作蛋白,但其互作仍需要BiFC等技术进一步验证。此外,CHCHD2的卷曲螺旋结构域是否为p26的直接结合区域,以及该结构域突变是否影响互作特异性,需通过结构域截断和定点突变实验明确。后续为解析CHCHD2的功能,可以通过病毒诱导的基因沉默(VIGS)或CRISPR-Cas9技术敲低CHCHD2,观察APV1病毒载量变化,明确其在病毒侵染中的功能。CHCHD2作为连接病毒侵染与宿主免疫的枢纽分子,其功能解析将为槟榔黄化病的防控提供新的靶标。这些发现为阐明APV1的致病机制提供了分子基础,同时为槟榔抗病品种选育提供了潜在靶点。未来结合功能基因组学与分子生物学技术,将进一步推动槟榔黄化病的综合防控策略开发。
Screening of host interaction proteins of areca palm velarivirus 1 (APV1) p26 via the yeast two-hybrid system
DOI: 10.15886/j.cnki.rdswxb.20250066
- Received Date: 2025-05-22
- Rev Recd Date: 2025-08-01
- Available Online: 2026-01-15
- Publish Date: 2026-01-25
-
Key words:
- areca palm velarivirus 1 (APV1) /
- p26 /
- yeast two-hybrid /
- host interaction proteins /
- CHCHD2
Abstract: Areca Palm Yellow Leaf Disease, caused by areca palm velarivirus 1 (APV1), poses a severe threat to the areca palm (Areca catechu L.) industry in Hainan. The p26 protein encoded by APV1 plays a critical role in viral infection, yet its interacting host proteins remain unidentified. The yeast two-hybrid (Y2H) technique was employed to screen an Areca palm cDNA library using p26 as the bait to identify interaction partners. The pGBKT7-p26 bait vector was successfully constructed and passed toxicity and autoactivation assays, confirming its suitability for the interaction screening system. Six candidate interacting proteins were identified by library screening, including Ras-related protein RABE1c, chloroplast ferritin-4 (Ferritin-4), oil palm stress-responsive protein PHOS34, and three proteins of unknown function. Following secondary screening and sequencing alignment, the focus was converged on an uncharacterized protein harboring a coiled-coil domain—coiled-coil helix-coiled-coil helix domain-containing 2 (CHCHD2). Retransformation assays confirmed the specific interaction between p26 and CHCHD2 in yeast. Bioinformatics analysis revealed that CHCHD2 has a molecular formula of C628H1007N201O198S8 and a molecular weight of 14.8 kDa. It is predicted to be a hydrophilic, unstable protein, with its secondary structure dominated by random coils. Subcellular localization predictions target CHCHD2 to the chloroplast, and it lacks signal peptides or transmembrane domains. This study reveals an in vitro interaction between APV1 p26 and the host protein CHCHD2, laying the groundwork for elucidating the function of p26 in viral pathogenesis and the CHCHD2-mediated host defense mechanisms.
| Citation: | Liang Yongqi, Wang Yutian, Gao Baosen, Zhao Ruibai, Wang Hongxing, Li Jin, Cao Xianmei. Screening of host interaction proteins of areca palm velarivirus 1 (APV1) p26 via the yeast two-hybrid system[J]. Journal of Tropical Biology, 2026, 17(1): 91-100. doi: 10.15886/j.cnki.rdswxb.20250066

|
 DownLoad:
DownLoad: