
-
莠去津(Atrazine)又名阿特拉津,分子式为C8H14ClN5,属于三氮苯类化合物[1-2]。莠去津通过干扰光合系统Ⅱ上的电子流动,破坏细胞膜完整性,最终导致杂草植株死亡[3],主要用于防除1年生禾本科杂草和阔叶杂草,对某些多年生杂草也有一定的抑制作用[4]。莠去津自1959年投入商业生产以来在全世界范围内广泛施用[5],占美国全年除草剂使用量的60%[6]。在中国华北和东北地区,莠去津及其混剂也是应用最广泛的除草剂[7]。莠去津属于低毒类除草剂,长期接触可对生物机体产生亚急性损伤[8-9]。莠去津使用后残留时间较长,水中的半衰期为15~20 d,在土壤中的半衰期可达330~385 d[10]。近年来,莠去津因其使用量大、土壤残留期长、环境中检出率高及潜在的内分泌干扰效应等受到广泛关注[11-12]。
在土壤和水体生态系统中,莠去津的代谢途径主要分为化学降解和生物降解。化学降解包括光解、水解、氧化-还原降解等。而生物降解途径则依赖于细菌、真菌和藻类等多种微生物类群的作用[13]。目前已知的莠去津降解细菌主要属于假单胞菌属(Pseudomonas)[14]、红球菌属(Rhodocdcus)[15]、不动杆菌属(Acinetabacter)、土壤杆菌属(Agrobacterium)[16]、蜡状芽孢杆菌(Bacillus cereus)[17]、诺卡氏菌属(Nocardia)[18]等。真菌属主要有曲霉属(Aspergillus)、青霉属(Penicillium)、木霉属(Trichoderma)、镰刀菌属(Fusarium)[19-20];藻类有衣绿藻属(Chlamydomonas)等[20]。
木霉(Trichoderma spp.)是农业生产中应用广泛的一种重要生防菌,同时还具有降解多种环境污染物的能力,可用于修复农化物质对环境的污染[21-22]。王素娜等[23]利用莠去津降解木霉菌株Ttrm11和Ttrm12对莠去津残留土壤进行修复研究。祁金玉等[24]探究了绿木霉(T. virens)和哈茨木霉(T. harzianum)在不同浓度阿特拉津处理下的生长速率、生物量和酶活特性。关于如何提升木霉降解莠去津的能力却鲜有报道, 因此,本研究通过含药培养基筛选出一株降解莠去津木霉菌株HN154,并通过单因素试验和响应面法优化培养条件,确定菌株HN154降解莠去津的最佳培养基配方,为后续莠去津的微生物降解提供理论基础。
-
从常年施莠去津的土壤里分离出木霉菌株HN154、AH003、AH009、FJ002、HL008、HL201、FJ059、LS029-3、SC106和WL043-2,均为海南大学热带农林学院植物病害微生态学研究与防控团队分离并保存。PDA培养基配方[25]:马铃薯 0.2 kg·L−1,无水葡萄糖 0.02 kg·L−1,另加琼脂粉 20.0 g·L−1。
Burk培养基配方[25]:KH2PO4 800.0 mg·L−1,K2HPO4 200.0 mg·L−1,CaSO4·2H2O 100.0 mg·L−1,MgSO4·7H2O 200.0 mg·L−1,(NH4)2SO4
1000.0 mg·L−1,MnSO4 10.0 mg·L−1,FeSO4·7H2O 5.0 mg·L−1,Na2MoO4·2H2O 3.3 mg·L−1,无水葡萄糖100.0 g·L−1。以莠去津为唯一碳氮源时,不添加(NH4)2SO4和无水葡萄糖;配制固体培养基时,添加琼脂粉20.0 g·L−1。 -
供试大豆品种为‘中黄37’。莠去津原药(98%)购于阿拉丁公司。色谱级的甲醇和二氯甲烷购自阿拉丁公司。其他常见分析纯试剂和药品购自白鲨易(biosharp)。
-
参考孙佳楠等[26]的试验方法,将木霉菌株分别接种于PDA平板,随后放置在28 ℃的恒温环境中活化,每隔3 d实施1次传代,待完成3次传代培养后,菌株供后续实验。
使用无菌水洗脱孢子,吸取1 mL孢子悬液(2×109 CFU·mL−1),接种至200 mL PD培养基中,28 ℃、180 r·min−1,振荡培养48 h后,利用真空抽滤装置对培养液进行抽滤处理,无菌水冲洗菌丝,再将其压干备用。
-
将木霉菌株分别接种至PDA培养基平板上,28 ℃培养96 h后,用打孔器打取6 mm菌饼接种到以
5000 mg·L−1莠去津为唯一碳氮源的无机盐培养基平板中,28 ℃倒置培养,96 h内观察记录木霉生长速率。通过比较菌丝生长速率筛选出对莠去津耐受效果最优菌株进行后续实验。将莠去津添加到Burk培养基平板中,使莠去津最终浓度分别为0、100、250、500、750、
1000 、1500 mg·L−1,取培养96 h的木霉平板,用打孔器打取6 mm菌饼,分别接种到含不同浓度莠去津的培养基中央,持续观察并记录各平板上木霉的菌丝生长直径,用十字交叉法测量菌落直径,制作菌落直径生长曲线,每个处理重复5次[27]。 -
将菌丝用滤纸过滤,称取0.5 g菌丝,121 ℃、30 min灭菌。将灭菌的木霉菌丝体(HN154-heatkilled)与未灭菌的木霉菌丝体(HN154-Live)分别加入到50 mL以500 mg·L−1莠去津为唯一碳氮源的Burk培养基中,28 ℃、180 r·min−1摇培72 h,在获得的培养液中分别加入等体积二氯甲烷振荡萃取30 min,静置分层后收集上层二氯甲烷相,重复3次。合并萃取液,加入1 g无水硫酸镁除水,旋蒸干燥(水温≤30 ℃),用色谱级甲醇溶解,经0.22 μm滤膜过滤。样品经高效液相色谱仪进行检测,检测条件为流动相为水-甲醇(V:V = 50:50),流速1 mL·min−1,检测波长220 nm,进样量20 μL,柱温30 ℃,液相色谱样品制备及检测条件参考朱云鹏等[28]。
莠去津样品标准曲线的制作:用色谱甲醇把莠去津标准溶液逐步精准稀释成 500、400、300、200、100、80、60、40、20 mg·L−1 质量浓度系列,依次用高效液相色谱仪检测。以峰面积为纵坐标,农药含量为横坐标绘制标准曲线,重复实验5次。
-
将HN154接种在PDA培养基上,26 ℃黑暗培养,每隔24 h观察菌株形态、大小、颜色、表面、边缘及生长速率等特征。3 d后,挑取菌丝于光学显微镜下观察并拍照记录[29]。
-
提取HN154脱氧核糖核酸(DNA),使用引物ITS1(5′-GCACAATAAGCGGAGGAAAAG-3′)和ITS4(5′-GGTCCGTGT-TTCAAGACGG-3′)[30]扩增内转录间隔区(ITS)的基因序列;利用对应的通用引物 PR2-F(序列为 5′-CGCCAGGGTTTTCCCAGTCACGACATGGCCTACATGAAGCGAT-3′)与 PR2-R(序列为 5′-AGCGGATAACAATTTCACACAGGAATCTCACAATGCGTGTACATGT-3′)扩增编码RNA聚合酶Ⅱ第2大亚基(RPB2)的基因序列。进一步测序和Blast(
https://blast.ncbi.nlm.nih.gov/Blast.cgi )分析鉴定,利用MEGA v11软件构建系统树。通过亲缘关系分析确定分离株的种属分类[31]。 -
将HN154接种至PDA培养基平板中,28 ℃培养96 h,用20 mL ddH2O将平板中木霉孢子洗脱,稀释孢子浓度至1×107 CFU·mL−1备用。以莠去津降解率为评价指标,依次探究葡萄糖质量浓度(0、10、15、20、25 g·L−1)、初始pH(5、6、7、8、9)、接种量(0.5%、1.0%、1.5%、2.0%、2.5%)和培养温度(20、25、30、35 ℃)4个因素对HN154降解莠去津的影响。实验重复5次。
-
利用Design-Expert软件设计4因素3水平响应面优化试验。根据单因素实验数据进行回归分析,得到以莠去津降解率为参考值的回归模型。利用该模型对HN154的发酵条件进行实验和分析。重复预测试验,取平均值,以验证模型是否可靠,进而得出最终优化结果。实验方法参考耿凯伟等[32]。分析方法参考王珊珊等[33]。
-
采用单因素方差分析(ANOVA)与Tukey检验评估组间统计差异,每处理设5次重复,结果用“平均值±标准差”表示。数据利用Design-Expert v10、SPSS v26.0和OriginPro v9.5软件处理分析。
-
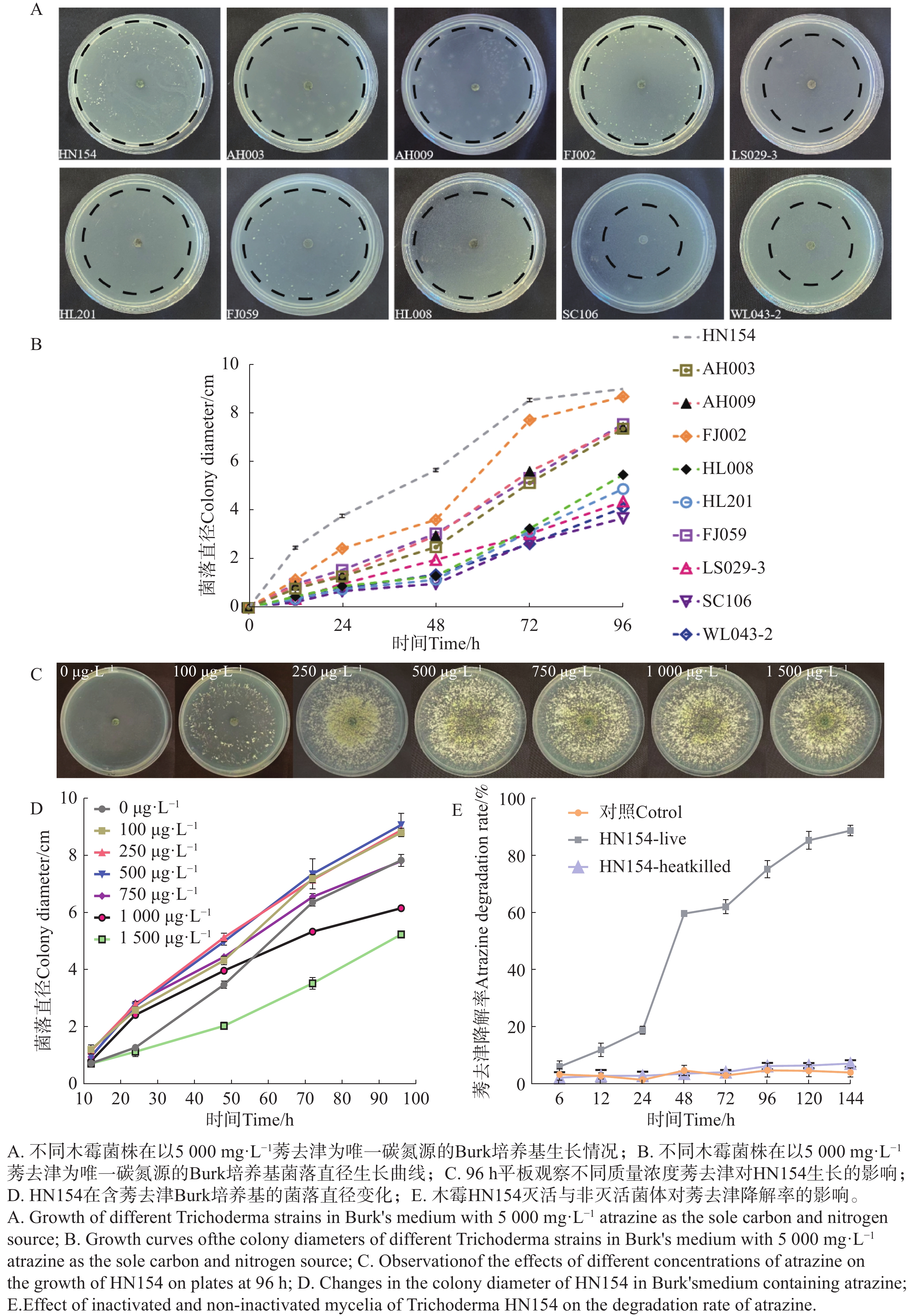
通过以高质量浓度莠去津为唯一碳源的Burk培养基,对10株木霉菌株进行耐药性测定,结果(图1-A—B)显示,在平板筛选结果中菌株HN154在
5000 mg·L−1莠去津为唯一碳氮源的Burk培养基中生长状况最佳,72 h菌丝即可覆满平板,且后续产孢能力显著优于其他菌株。菌株FJ002需培养96 h可覆满平板;其余菌株菌丝生长缓慢,培养96 h仍未能覆满平板。使用不同质量浓度梯度的莠去津平板,测定莠去津对菌株HN154生长的影响,结果(图1-C—D)显示,莠去津浓度为500 mg·L−1时HN154生长最快,96 h菌丝布满平板且产孢浓密。为了进一步鉴定HN154降解莠去津的能力,将未灭活和灭活的HN154菌丝体放在以500 mg·L−1莠去津为唯一碳氮源的Burk培养基中培养,结果显示灭活的莠去津降解速率与对照无显著性差异,不能降解莠去津,非灭活的莠去津降解速率在24~48 h从20.3%上升至60.5%,呈现指数上升趋势,整体降解率随时间延长而显著提高,144 h降解率达到90.1%,表明HN154具有降解莠去津能力(图1-E)。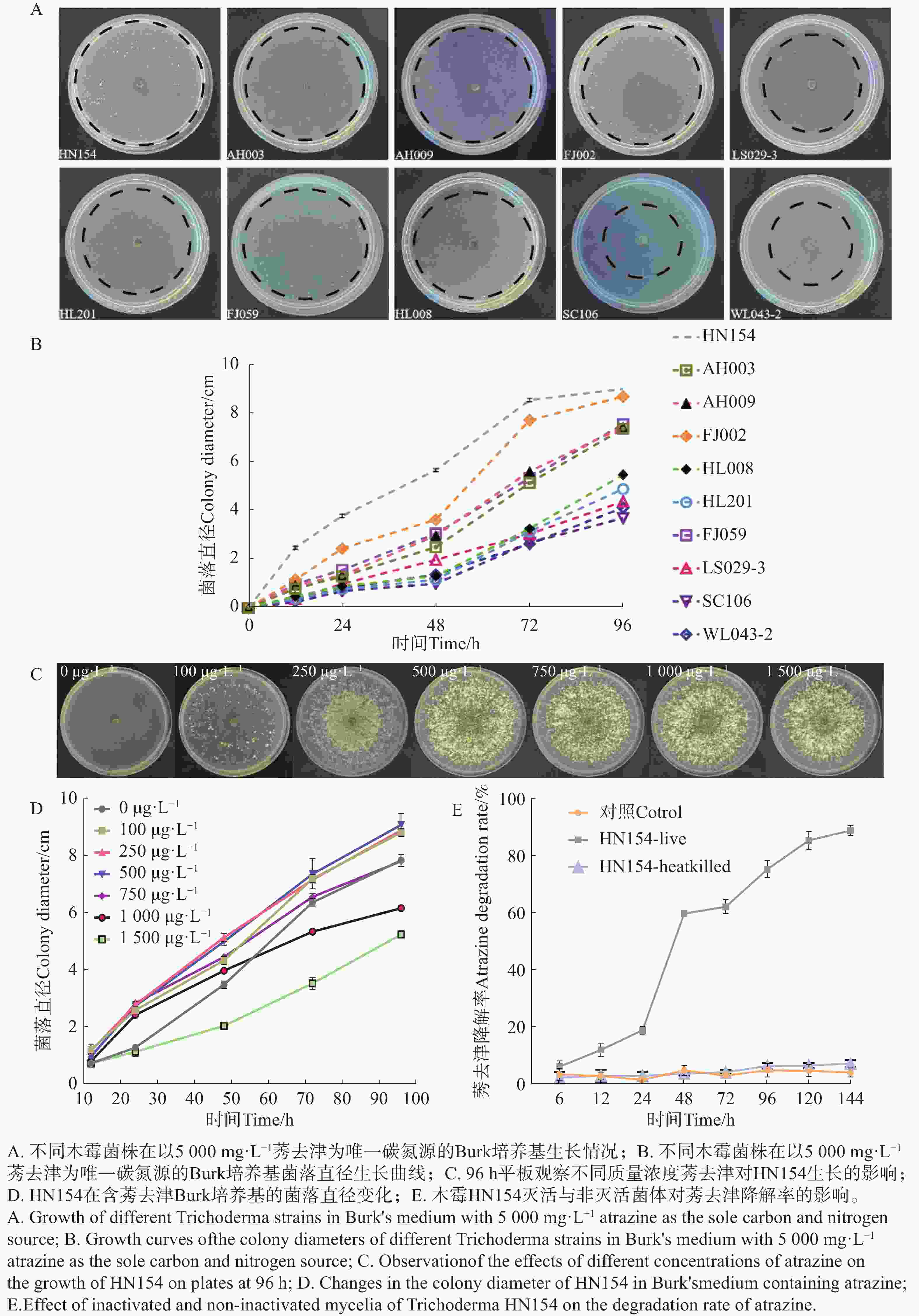
Figure 1. Screening of atrazine-degrading Trichoderma strain
-
HN154菌落呈同心圆形,初期气生菌丝呈白色,羊毛状,菌丝交错,在菌丝生长后期产生分生孢子,分生孢子深绿色,孢子簇棉絮状,分布在菌丝密集处;分生孢子梗常呈现2~3个孢子梗分支,顶端具有多个瓶梗,瓶梗与主轴夹角约75°距顶端较远处产生至多三级分枝;瓶梗产生于分枝顶端,3~5个呈漩涡状排列,瓶梗直、中间稍有加粗,顶部、基部缢缩;分生孢子球形、近圆形(图2)。
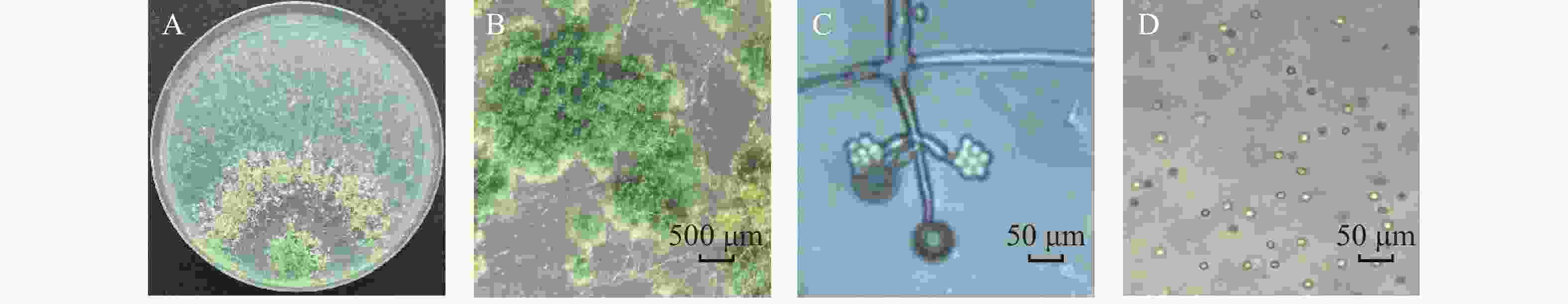
Figure 2. Morphological characteristics of strain HN154
-
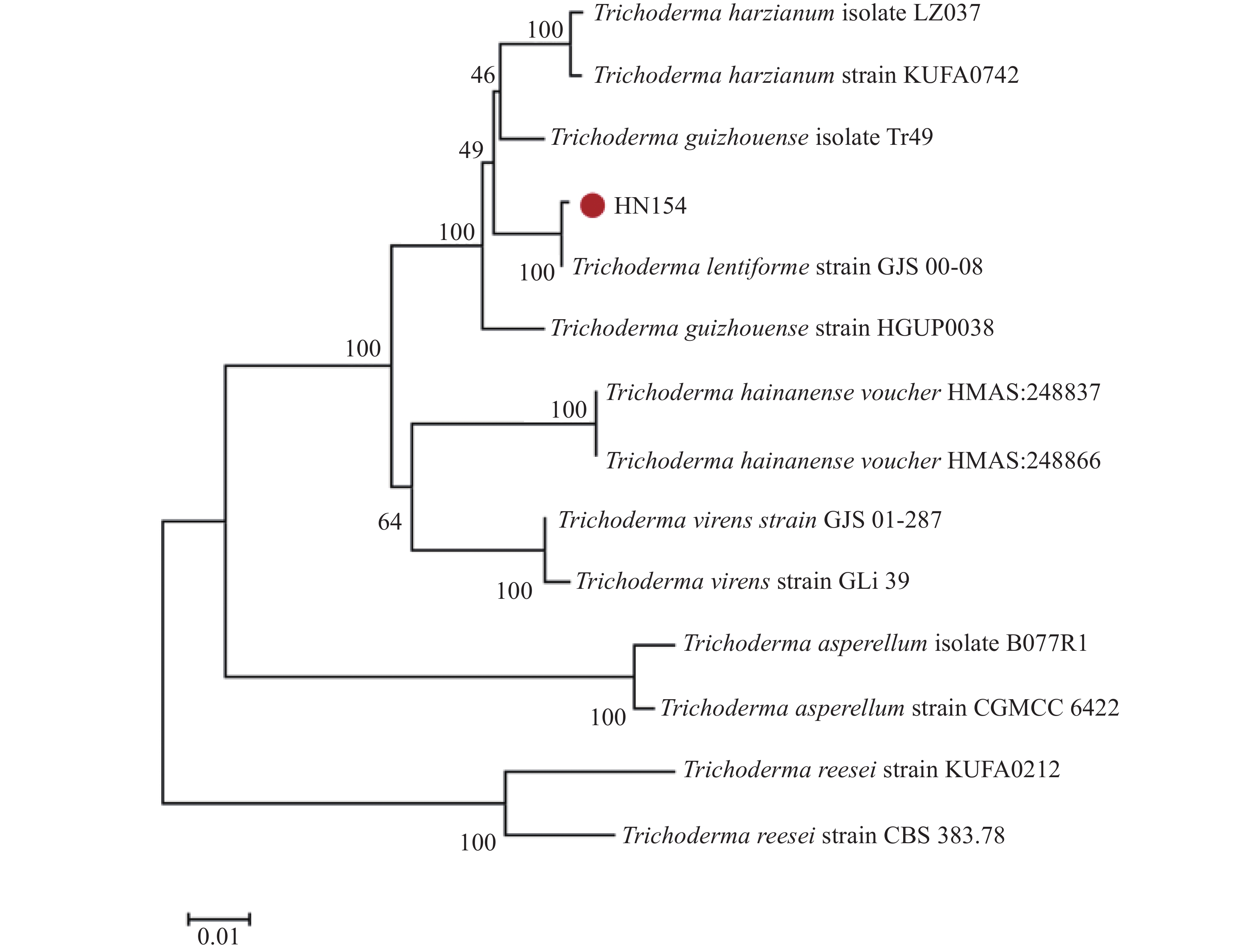
基于ITS与RPB2基因序列构建的系统树(图3)发现,菌株HN154和模式菌株Trichoderma lentiforme strain GJS00-08的亲缘关系最近,二者在系统树上分支距离极短,亲缘关系最近。结合形态学特征的观测结果,菌株HN154最终被鉴定为Trichoderma lentiforme。
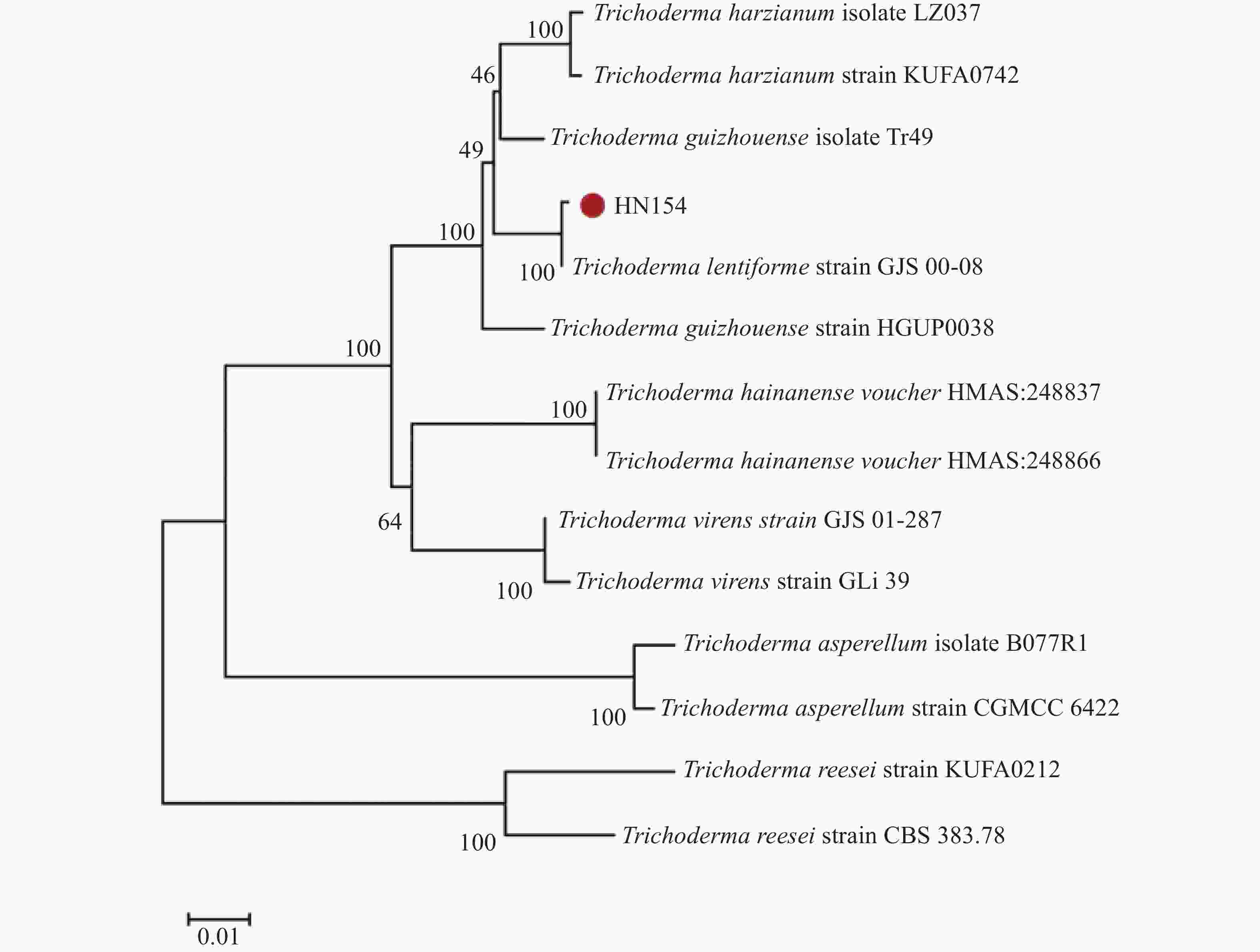
Figure 3. Phylogenetic tree of HN154 constructed based on ITS and RPB2 sequences
-
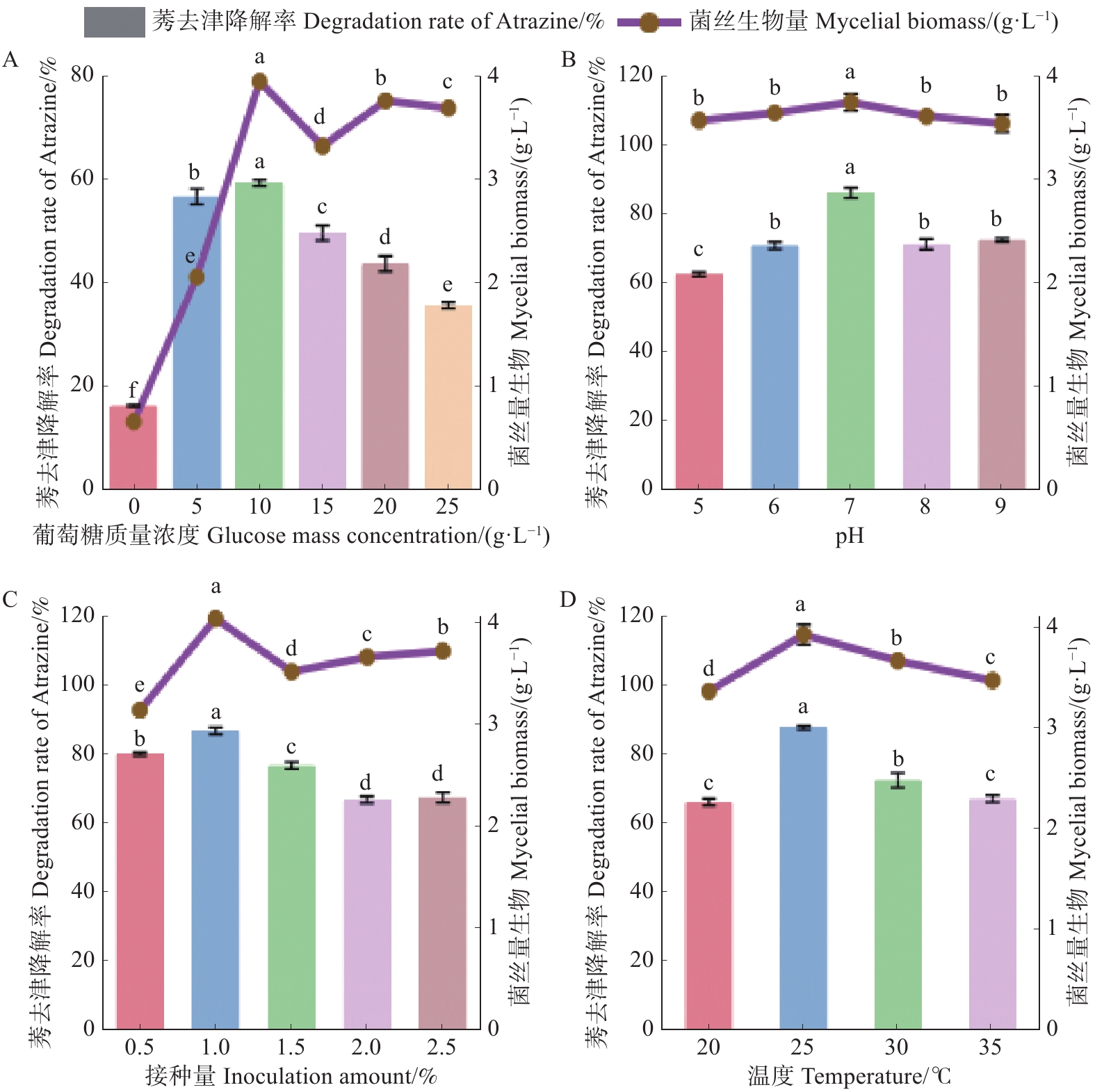
不同质量浓度葡萄糖的培养基对菌株HN154生长及降解莠去津能力的测定结果(图4-A)显示,菌株HN154对莠去津降解率随葡萄糖质量浓度呈先升后降的趋势。当葡萄糖质量浓度为10 g·L−1时,菌丝生物量最高,为3.85 g·L−1;其对莠去津的降解率也最高,为59.51%。
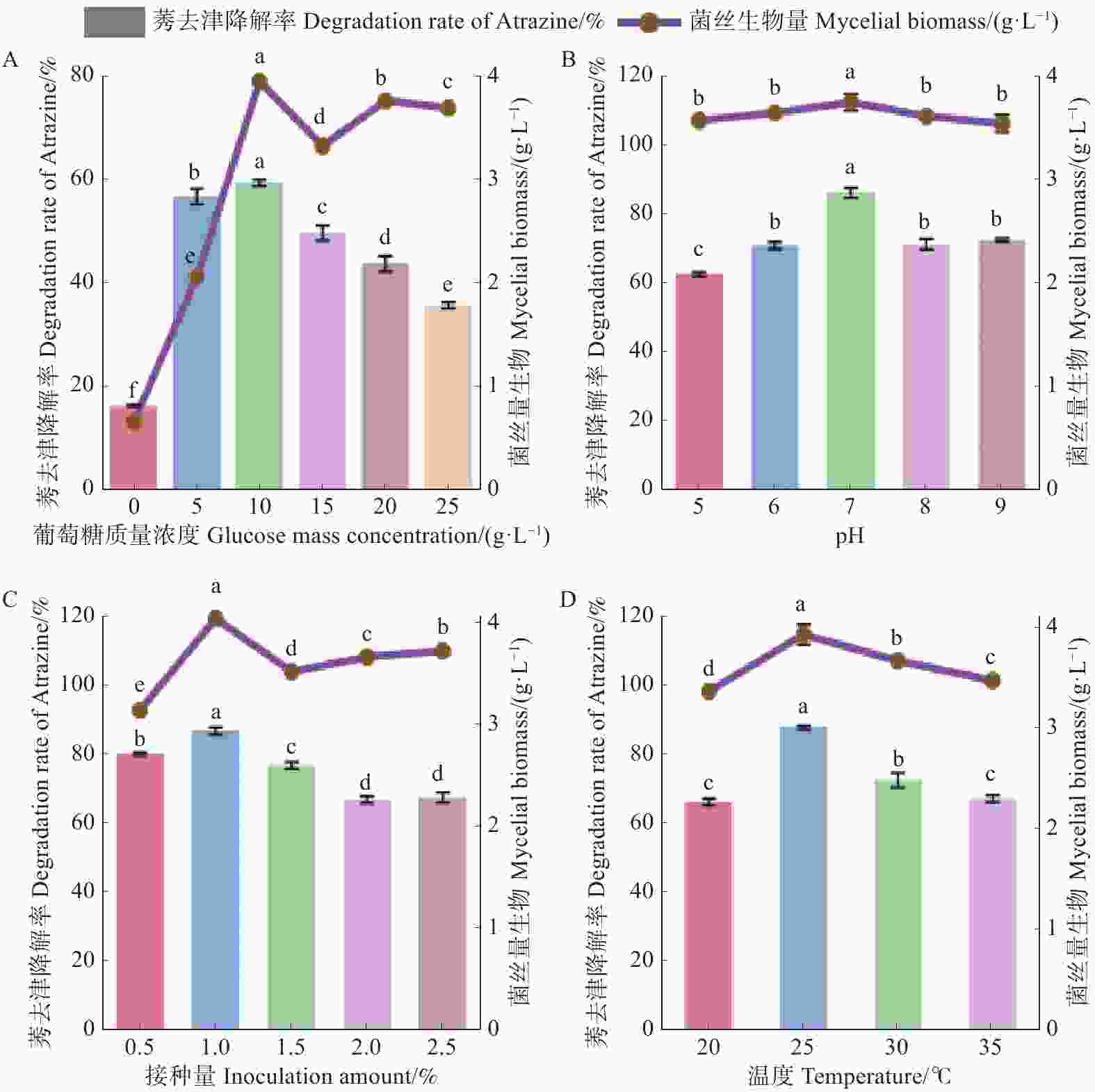
Figure 4. Effects of different conditions on the mycelial growth of HN154 and atrazine degradation
不同初始pH的培养基对菌株HN154生长及其降解莠去津能力的测定结果(图4-B)显示,菌株HN154对莠去津降解率随初始pH呈先升后降的趋势。当初始pH7时,菌丝生物量最高,为3.89 g·L−1;其对莠去津的降解率也最高,为86.75%。
不同接种量的培养基对菌株HN154生长及其降解莠去津能力的测定结果(图4-C)显示,菌株HN154对莠去津的降解率随木霉接种量呈先升后降的趋势。当接种量为1%时,菌丝生物量最高,为3.93 g·L−1;其对莠去津的降解率也最高,为87.62%。
不同发酵温度的培养基对菌株HN154生长及其降解莠去津能力的测定结果(图4-D)显示,菌株HN154对莠去津的降解率随发酵温度呈先升后降的趋势。当发酵温度为25 ℃时,菌丝生物量最高,为3.96 g·L−1;其对莠去津的降解率也最高,为87.79%。
-
依据单因素试验设计所获结论,针对4个自变量开展不同组合,总计进行29次试验,具体的试验设计与结果详见表1。借助预测并经过回归拟合处理,得出了以莠去津降解率(Y)作为响应值的回归方程:
实验号
Experimental No.a 温度/℃
Temperatureb pH c 接种量/%
Inoculum amountd 葡萄糖质量浓度/(g·L−1)
Glucose mass concentrationY莠去津降解率/%
Atrazine degradation rate1 40 7 1.5 25.0 75 2 30 7 2.5 0 77 3 30 9 1.5 0 79 4 20 7 2.5 12.5 71 5 30 7 1.5 12.5 83 6 30 7 0.5 0 69 7 20 7 0.5 12.5 70 8 30 5 2.5 12.5 71 9 30 5 1.5 25.0 82 10 20 5 1.5 12.5 70 11 40 5 1.5 12.5 75 12 40 9 1.5 12.5 75 13 30 9 0.5 12.5 72 14 30 7 1.5 12.5 83 15 40 7 0.5 12.5 75 16 30 7 1.5 12.5 83 17 30 7 1.5 12.5 82 18 30 9 2.5 12.5 76 19 30 7 2.5 25.0 75 20 30 9 1.5 25.0 70 21 30 5 0.5 12.5 75 22 30 7 1.5 12.5 84 23 40 7 2.5 12.5 75 24 20 7 1.5 0 69 25 30 7 0.5 25.0 77 26 20 7 1.5 25.0 76 27 30 5 1.5 0 65 28 20 9 1.5 12.5 70 29 40 7 1.5 0 75 Table 1. Response surface experimental design and results
Y=83.00+2.00a+0.33b+0.58c+1.75d+0.000ab−0.25ac−1.75ad+2.00bc−6.50bd−5.50a2−5.00b2−4.36c2−3.88d2
由表2可知,该模型的P值小于0.000 1,达到了极显著水平;失拟项的P值为0.260 5,大于0.05,未呈现出显著差异,这意味着模型具备较高的可靠性。变异系数(CV)仅为1.24%,处于较低水平,进一步佐证了模型的可信度。决定系数R2的数值是0.983 0,这表明模型与测试结果数据的符合程度高达98.30%。调整决定系数R2adj为0.965 9,说明此方程对试验的拟合效果良好。此外,由表2还可知,一次项中的a、c、d,交互项里的ad、bc、bd、cd,以及二次项的a2、b2、c2、d2,它们对结果的影响极为显著;其余各项对结果的影响并不显著(P<0.01)。
来源
Source平方和
Squares自由度
df均方
Mean squareF值
F ValueP值
Prob > FProb > F显著性
Significance模型Model 697.36 14 49.81 57.71 < 0.0001 ** a-温度Temperature 48.00 1 48.00 55.61 < 0.0001 ** b-pH 1.33 1 1.33 1.54 0.2343 c-接种量
Inoculation amount4.08 1 4.08 4.73 0.0473 ** d-葡萄糖浓度
Glucose concentration36.75 1 36.75 42.58 < 0.0001 ** Ab 0.000 1 0.000 0.000 1.0000 Ac 0.25 1 0.25 0.29 0.5989 Ad 12.25 1 12.25 14.19 0.0021 ** Bc 16.00 1 16.00 18.54 0.0007 ** Bd 169.00 1 169.00 195.81 < 0.0001 ** Cd 25.00 1 25.00 28.97 < 0.0001 ** a2 196.22 1 196.22 227.34 < 0.0001 ** b2 162.16 1 162.16 187.88 < 0.0001 ** c2 138.75 1 138.75 160.76 < 0.0001 ** d2 97.40 1 97.40 112.85 < 0.0001 ** 残差Residual 12.08 14 0.86 失拟项Lack of Fit 10.08 10 1.01 2.02 0.2605 变异系数CV 1.24 ** 净误差Net error 2.00 4 0.50 合计Total 709.45 28 决定系数R2 0.9830 adj决定系数Radj2 0.9659 注:**,P< 0.0001 。Note: **, P < 0.0001 .Table 2. ANOVA table of the regression model
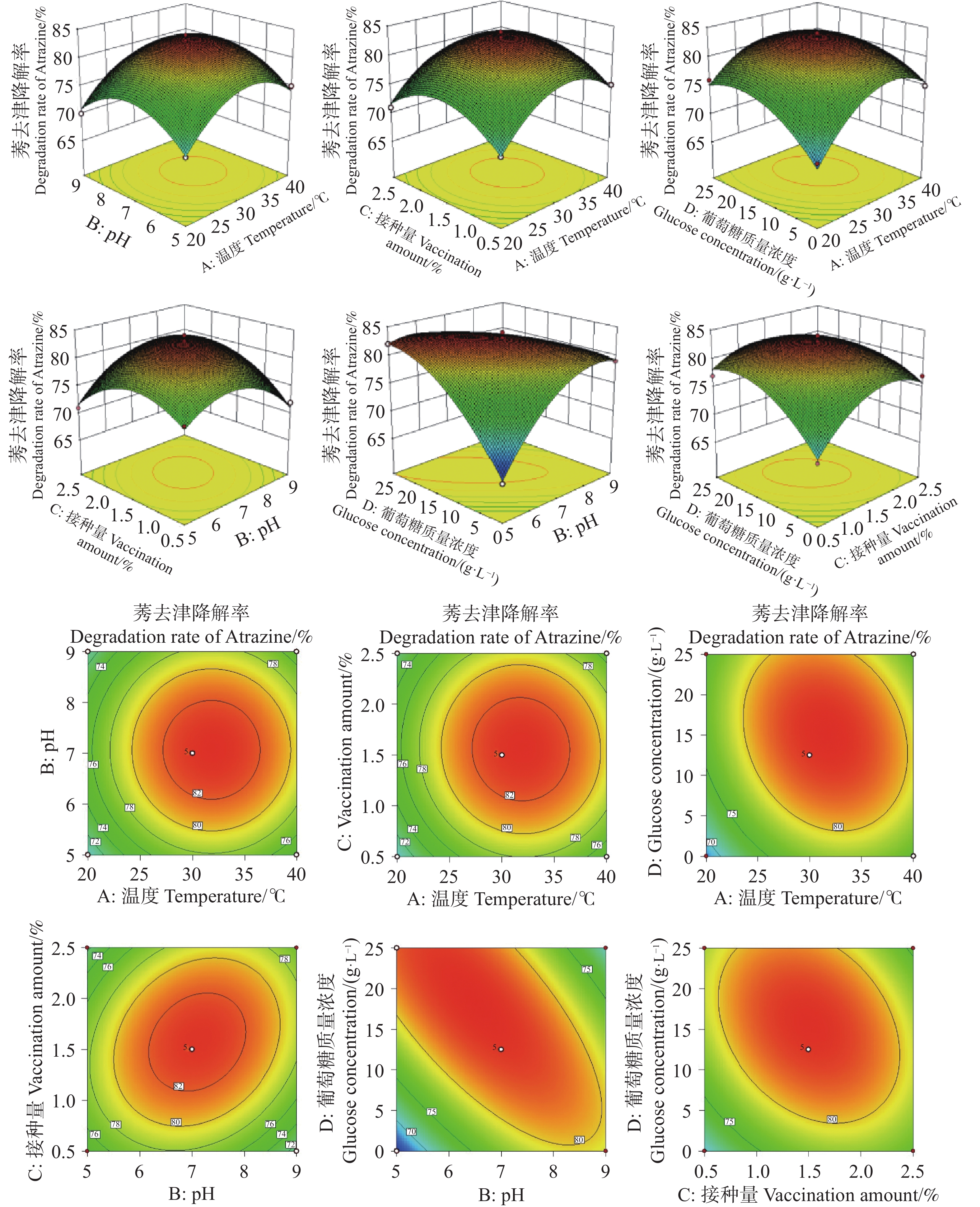
图5所示的响应面模型,直观且清晰地呈现出各因素之间的交互作用给HN154降解莠去津的影响。其中,3D响应面呈现出凸面形态,意味着在当前因素组合下,均存在对应的降解率最大值。在交互项中,ad、bc、bd、cd的等高线呈现为椭圆形,依据响应面分析的原理,表明这些交互项对莠去津降解率有着较大的影响力;与之相反,ab、ac的等高线形态则显示出它们对莠去津降解率的影响相对较小,与表2的方差分析结果相互印证。
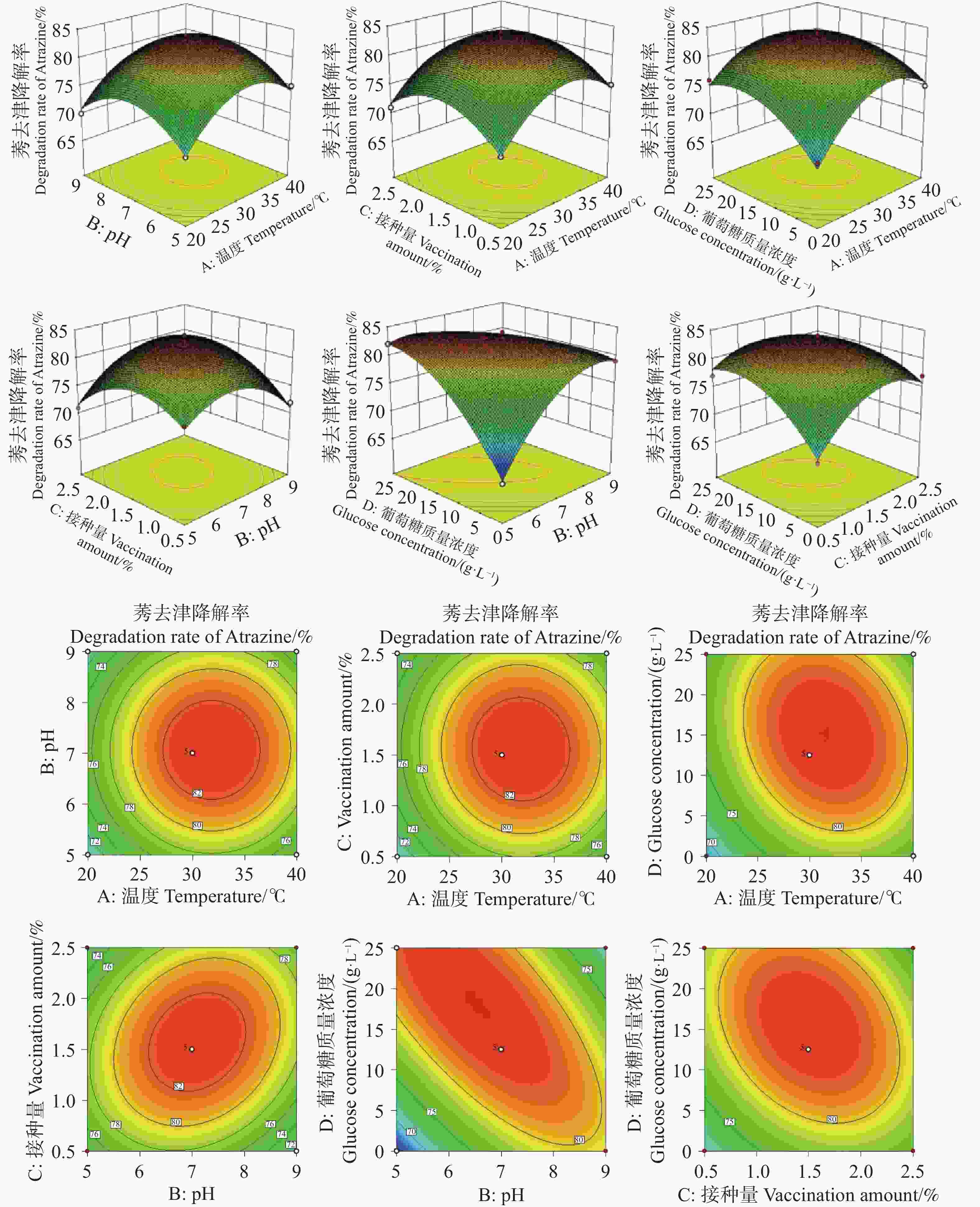
Figure 5. Three-dimensional response surface model and contour line model of the four-factor three-level design
通过回归模型预测莠去津降解率的最佳参数为葡萄糖质量浓度5.24 g·L−1,pH 7.78,接种量0.77%,温度25.5 ℃;莠去津的理论最大预测降解率为88.57%。应用回归模型预测莠去津降解率的最佳参数进行验证。HN154(T. lentiforme)在以500 mg·L莠去津为唯一碳氮源的无机盐培养基中培养3 d时莠去津降解率最高达到89.02%,与理论最大预测降解率符合。
-
本试验从不同土壤中分离的10株木霉,以莠去津为唯一碳氮源筛选出生长较好的菌株HN154。该菌株在高浓度
5000 mg·L−1莠去津浓度下72 h菌丝即可覆满平板,且后续产孢能力显著优于其他菌株。灭活菌丝体降解能力与空白组相近,无降解能力;未灭活菌丝体降解效果显著,12~48 h内降解速率快速提升,降解曲线符合酶促反应规律。500 mg·L−1莠去津浓度可能接近HN154降解酶的最适底物浓度,此时酶活性最高,菌株生长最活跃。与前人研究一致,木霉对三氮苯类除草剂有强耐受性和氧化还原酶活性[34-35]。微生物降解农药的效率受多种因素影响,主要包括温度、pH、微生物特性、碳源和氮源的存在及其浓度等[36-37]。有研究表明,通过调节培养基中的葡萄糖质量浓度、pH、发酵温度和光照时间等条件,可显著影响木霉的生长及其对病原菌的抑制作用[38-39]。如葡萄糖的质量浓度可直接影响木霉的生长和代谢[40]。环境pH是调控木霉生理活动的关键因素,通过影响细胞膜通透性和酶活性,进而影响营养物质的吸收和利用效率[41]。木霉对pH变化具有代谢应激能力,初始pH接近中性或弱碱性有利于其生长和代谢[42]。接种量是木霉发酵过程中的重要参数,过高或过低的接种量均会影响木霉生长和发酵效率[43-44]。此外,发酵温度也是调控微生物生长的重要因素,25 ℃被证实为木霉营养生长的最适温度[45]。本研究通过优化葡萄糖浓度、初始pH、接种量和发酵温度,确定了HN154菌株的最佳培养条件:初始pH7、葡萄糖质量浓度10 g·L−1、接种量1%、温度25 ℃,在此条件下对莠去津的降解效率达到最优。这一结果与已有文献报道的研究数据具有相似性,进一步验证了培养条件对木霉降解莠去津效率的显著影响。
响应面分析法(RSM)因其高效性和精准性,已广泛应用于培养基和发酵条件的优化研究。该方法通过局部试验构建连续变量的曲面模型,能够以最少的实验次数快速确定多因子系统中的最优条件,并识别关键影响因素[46]。例如,李洋[47]利用RSM优化草酸青霉菌(Penicillium oxalicum)JG的发酵培养基,以滤纸酶活(FPA)为响应值,最终使FPA活性提升至0.366 IU·mL−1。类似地,Lotfy等[48]采用RSM优化黑曲霉产柠檬酸的培养基,显著提高了发酵效率。本研究采用RSM优化HN154的降解条件,通过调节葡萄糖浓度、pH、接种量和发酵温度,寻求最佳降解效率。结果表明,葡萄糖浓度不宜过高,与袁辉林等[49]、陈晨等[50]研究结果一致,而pH、接种量和发酵温度的优化结果与木霉的常规培养条件相符[51]。最终优化条件为葡萄糖质量浓度5.24 g·L−1、pH7.78、接种量0.77%、温度25.5 ℃,在此条件下,HN154对500 mg·L−1莠去津的降解率提升至89.02%。目前,真菌降解莠去津的机制尚未完全明确。尽管如此,利用微生物降解莠去津以减轻莠去津污染的做法,因其成本低廉,处理效果显著,对环境影响小,且不会破坏植物生长所需的土壤环境等优点,仍得到了广泛的应用[52]。未来,运用微生物技术来降解莠去津并修复受污染的土壤和水体,将继续成为研究热点。
-
本研究成功筛选出高效降解莠去津的木霉菌株HN154,并通过优化培养条件显著提升了其降解效率,为农药残留的生物治理提供了具有应用潜力的真菌资源。实验结果为中国农业污染治理及生态安全维护提供了新的技术支撑与理论依据。未来可进一步探究HN154的降解代谢机制及田间应用潜力,以推动绿色农业与可持续发展目标的实现。
Screening and identification of atrazine-degrading trichoderma strains and optimization of degradation conditions
DOI: 10.15886/j.cnki.rdswxb.20240204
- Received Date: 2024-12-31
- Accepted Date: 2025-04-11
- Rev Recd Date: 2025-02-22
- Available Online: 2025-04-23
- Publish Date: 2026-05-25
-
Key words:
- atrazine /
- Trichoderma /
- response surface method /
- degradation of pesticide residues /
- weedicide
Abstract: The inorganic salt medium with atrazine as the sole carbon and nitrogen source was used to screen a Trichoderma strain with the best growth performance among 10 Trichoderma strains, and the Trichoderma strain HN154 was selected. The degradation test of atrazine demonstrated that this strain had a high ability to degrade atrazine. Strain HN154 was identified as Trichoderma lentiforme based on morphological characteristics and ITS/RPB2 gene sequence analysis. Optimization of atrazine degradation conditions showed that the atrazine degradation rate was as high as 87.79% when cultured in inorganic salt medium with 10 g·L−1 glucose, pH7, 25 ℃, and 1% inoculum. Response surface optimization further improved degradation to 89.02% under the conditions of 5.24 g·L−1 glucose, pH7.78, 0.77% inoculum, and 25.5 ℃. This study provides microbial resources and technical support for efficient atrazine degradation, offering insights into degradation of microbial pesticide residues.
| Citation: | Jiang Haonan, Lyu Yudie, Wu Yiping, Wu Guoliang, Wang Rui, Zhang Manman, Liu Tong. Screening and identification of atrazine-degrading trichoderma strains and optimization of degradation conditions[J]. Journal of Tropical Biology, 2026, 17(3): 451-462. doi: 10.15886/j.cnki.rdswxb.20240204

|
 DownLoad:
DownLoad: