-
主持人:廖卫国
由咖啡驼孢锈菌(Hemileia vastratrix)寄生引起的咖啡叶锈病在世界咖啡种植区普遍发生,是导致咖啡落叶和减产的关键病害,流行年份咖啡落叶率高达50%以上,减产率可达30%~50%,导致的经济损失高达10亿~20亿美元[1-3]。该病于1861年在非洲维多利亚湖畔首次发现。云南省于1956年开始推广种植小粒咖啡Coffea arabica,同年就发现了咖啡叶锈病[4]。病原菌H. vastratrix以锈孢子进行无性繁殖,而有性繁殖阶段至今未明确[5]。国内外学者对咖啡叶锈病的研究主要集中在农业景观生态系统及气候变化对咖啡叶锈病的影响[6-7]、危害特点、替代寄主植物[8]、生理小种[9]、化学防控[10]以及重寄生真菌[3, 11]等方面。咖啡产业作为云南省乡村振兴的重要产业,2020年全省种植咖啡面积8.91万ha,产量13.51万t,农业产值24.46亿元,是我国最大的小粒咖啡种植区,而干热区保山市则是云南省小粒咖啡主产区之一,咖啡种植面积8 741 ha,产量2.222 5万t,产值40 227.25万元[4]。自1956年以来,咖啡叶锈病一直是危害该地区的重要病害。海拔作为一个综合性生态因子,会引起光、温、水以及土壤等因子的综合变化,进而影响病虫害,但至今没有开展过不同海拔梯度对咖啡叶锈病及其重寄生菌影响的研究。基于此,本课题组通过为期1年对干热区咖啡叶锈病及其重寄生菌的发生动态进行监测,以期揭示海拔梯度对咖啡叶锈病及其重寄生菌的影响,进而为干热区不同海拔梯度咖啡园咖啡叶锈病的精准预测及防控策略的制定提供科学依据,也为咖啡叶锈病的生物防控提供重要数据支撑。
HTML
-
研究区域位于云南省保山市隆阳区潞江镇(25°1′N,98°49′E),属典型的干热河谷气候类型,具有全年旱雨季分明,降雨量低,昼夜温差大,年均温差小,全年无霜的特点。小粒咖啡多种植于怒江西部与高黎贡山延伸区间坡度小于35°、海拔600~1 800 m的区域。该区域海拔900 m以下土壤多为黑色沙土,海拔900 m以上多为红色壤土,偏中性或微酸性。该区域种植的咖啡品种以Catimor系列品种为主,部分区域种植少量的‘Typica’、‘Buorbon’、‘Geisha’等品种。该区域种植咖啡超过50 年。2014年前咖啡种植多以阳光咖啡种植为主,2014年后为提高产值,开始在咖啡园大面积套作杧果(Mangifera indica)、龙眼(Dimocarpus longan)、香蕉(Musa nana)等经济作物。
-
2020年10月至2021年9月,对云南省保山市潞江镇新寨村(25°1′35″N,98°49′51″E)受咖啡叶锈病侵染的咖啡植株进行观察和记录。
-
2020年10月至2021年9月,通过定点、定期对云南省保山市隆阳区新寨村、赧亢村、丛岗村的咖啡园咖啡叶锈病的周年动态进行监测。于每月的10日分别监测1次,每个监测点随机选择10株咖啡植株,从植株的上中下3个部位,分别选取10片咖啡叶片,使用数码相机Canon DS126271对采集的叶片进行拍照,测量咖啡叶锈病危害面积,计算危害面积百分比。
-
咖啡叶锈病周年动态监测同期,在云南省保山市隆阳区新寨村海拔700~1 500 m的区域,海拔每上升100 m选择1个咖啡园,共选9个咖啡园。每个咖啡园随机选择10株咖啡植株,每株于上中下3个部位,每个部位选择10片咖啡叶片,测量咖啡叶锈病危害面积,测量方法参照1.3。
-
在咖啡锈病危害高峰期(12月),于9个海拔梯度的咖啡园内随机选择10株咖啡植株,从每株咖啡植株的上中下3个部位,每个部位选择10片被咖啡叶锈病侵染的叶片,记录咖啡叶锈病是否被重寄真菌寄生,计算重寄生率。重寄生率=重寄生真菌寄生的叶片/总调查叶片×100%。
-
使用Excel 2021、SPSS 19.0、Origin 2021和Photoshop 2021等软件进行数据统计分析及绘图。
-
在干热区,保山咖啡叶锈病全年均有发生。病原菌H. vastratrix以锈孢子从咖啡叶片背面气孔侵入,危害初期在叶背可见2~3 mm黄色小斑,随着小斑的扩大,叶背产生肉眼可见的黄色或橙黄色孢子堆,随之孢子堆以同心圆的方式不断扩大,一般直径可达5~8 mm,后期多个病斑相互连接形成更大的病斑。初期叶正面则形成黄色圆形病斑,随着病斑老化中间出现褐色斑块。危害后期,整片叶片黄化、脱落,严重影响次年咖啡的产量。
-
在干热区,保山咖啡叶锈病周年动态具有3个高峰期,分别在12月和翌年的2月、4月,各代峰距约为60 d,以12月危害最严重,3个高峰期咖啡叶锈病的危害百分比分别为(4.12±0.66)%、(3.53±0.62)%和(2.38±0.42)%(图1)。

Figure 1. Annual damage dynamics of coffee leaf rust in dry-hot region
-
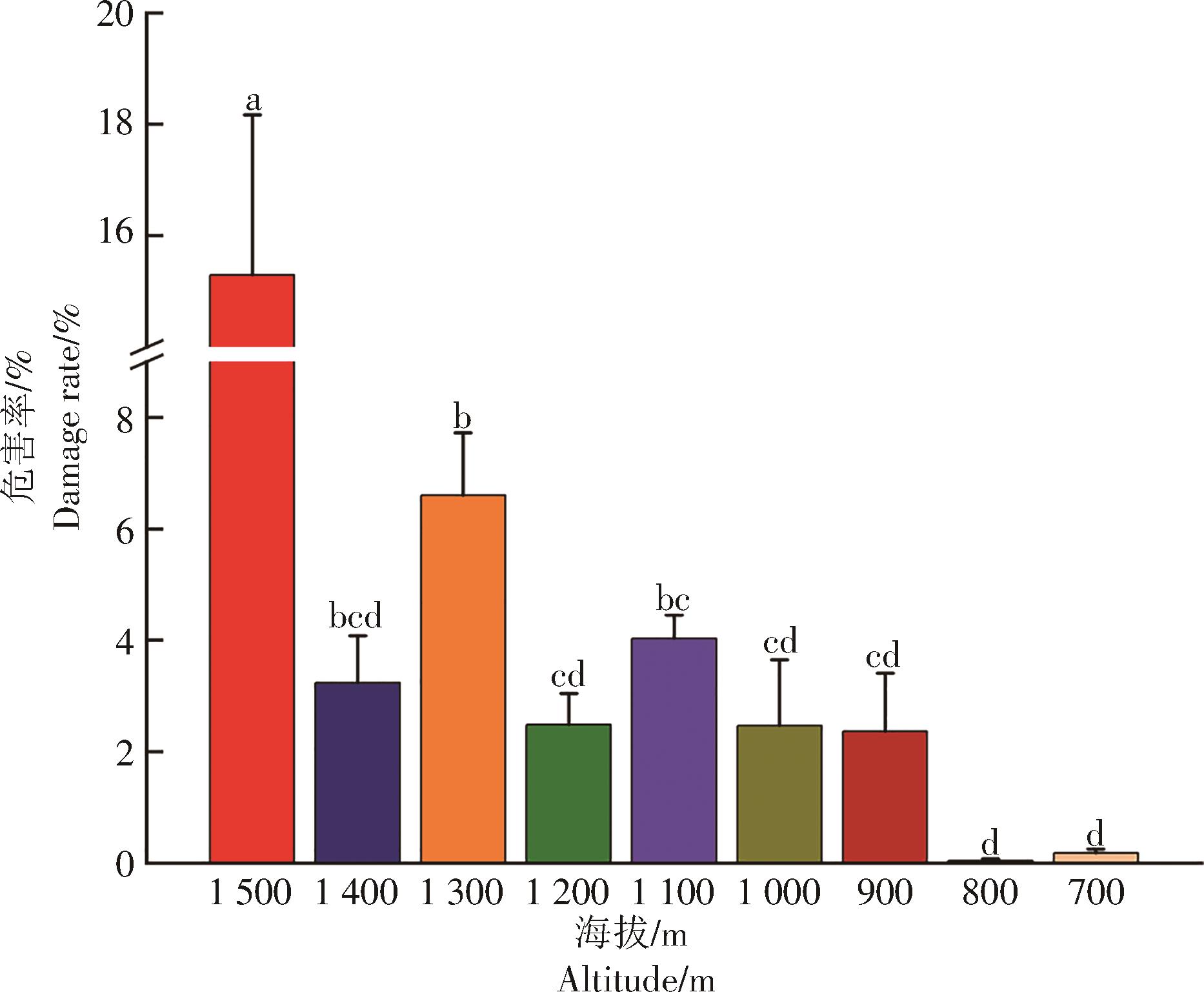
在干热区保山不同海拔梯度上咖啡叶锈病的危害百分比有差异(图2)。海拔1 500 m的咖啡叶锈病危害百分比显著高于其他8个海拔梯度的;海拔1 300 m的咖啡叶锈病危害百分比次之,显著高于1 200 m和1 000 m及其以下4个海拔梯度的,但与1 400 m和1 100 m的差异不显著;1 400 m和900~1 200 m的5个海拔梯度咖啡叶锈病的危害百分比差异不显著,不同海拔的危害百分比排序为1 100 m>1 400 m>1 200 m>1 000 m>900 m;低海拔700 m和800 m的咖啡叶锈病危害百分比低于1%,但700~1 000 m间4个海拔梯度咖啡叶锈病危害百分比差异不显著。总体来看高海拔地区咖啡叶锈病危害明显高于低海拔的。

Figure 2. Damage of coffee leaf rust at different altitudes
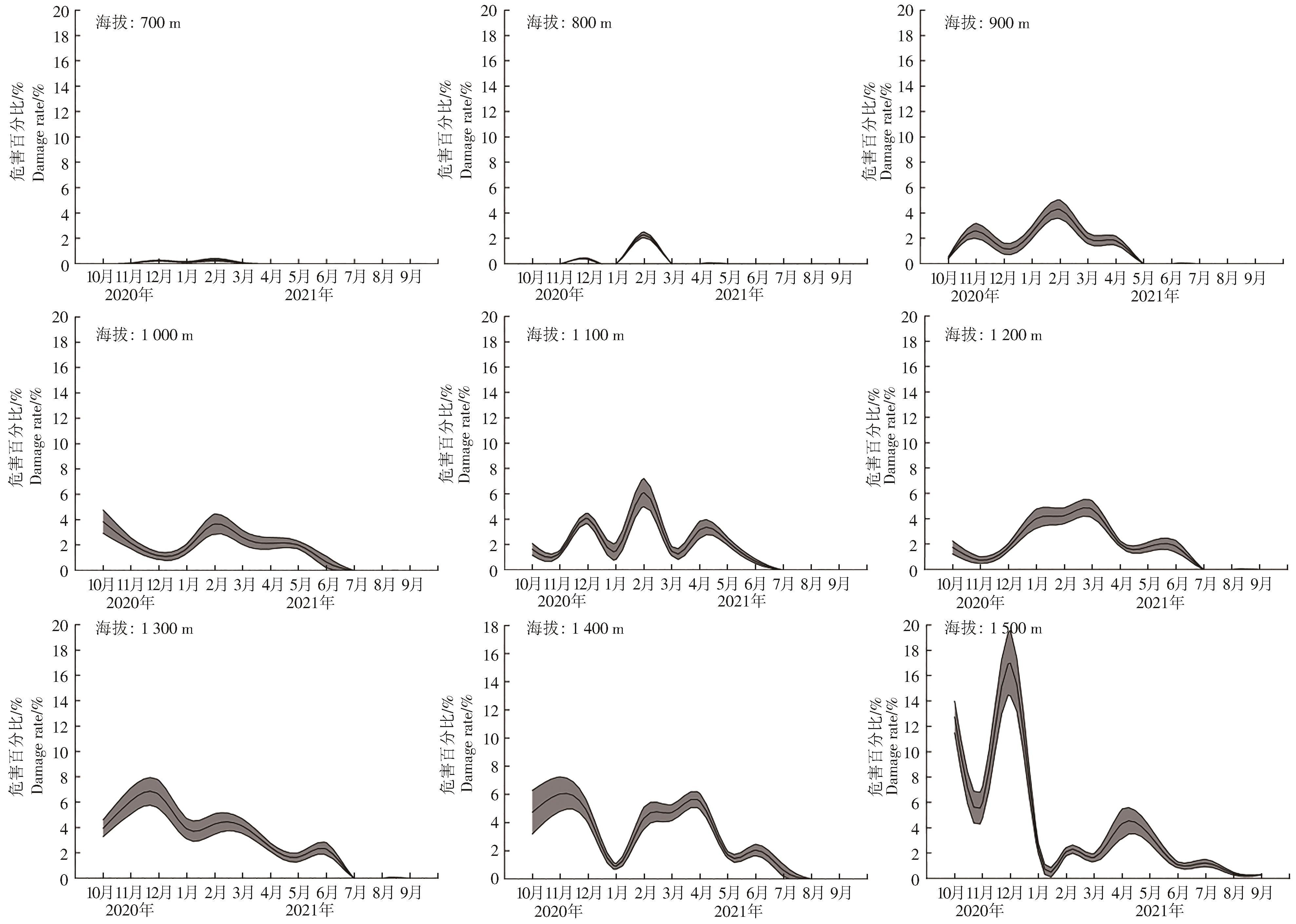
9个海拔梯度上的咖啡叶锈病周年变化规律不同(图3)。海拔700 m和800 m,咖啡叶锈病危害率均低于4%,仅12月和翌年2月发生;海拔900 m,咖啡叶锈病主要发生期为10月至翌年4月,具2个高峰,即11月和翌年2月,危害百分比分别为(2.59±0.60)%和(4.29±0.74)%;海拔1 000 m,7—9月咖啡叶锈病不发生,发生期仅有1个高峰,即翌年2月,为(3.66±0.80)%;海拔1 100 m,7—9月咖啡叶锈病不发生,发生期具3个高峰,分别为12月和翌年2月、4月,翌年2月咖啡叶锈病危害最高,为(6.09±1.11)%;海拔1 200 m,咖啡叶锈病7—9月不发生,发生期具2个高峰,分别为3月和6月,3月危害最高,为(4.74±0.68)%;海拔1 300 m,咖啡叶锈病7—9月不发生,发生期具3个高峰,分别为12月和翌年2月、6月,其中12月危害最严重,为(6.60±1.11)%;海拔1 400 m,咖啡叶锈病8月和9月不发生,发生期具3个高峰,分别为11月和翌年3月、6月,其中11月发生最严重,为(6.04±1.22)%;海拔1 500 m,咖啡叶锈病全年发生,具4个高峰,分别为12月和翌年2月、4月、7月,其中12月发生最严重,为(16.99±2.60)%。

Figure 3. Annual damage dynamics of coffee leaf rust at different altitudes
-
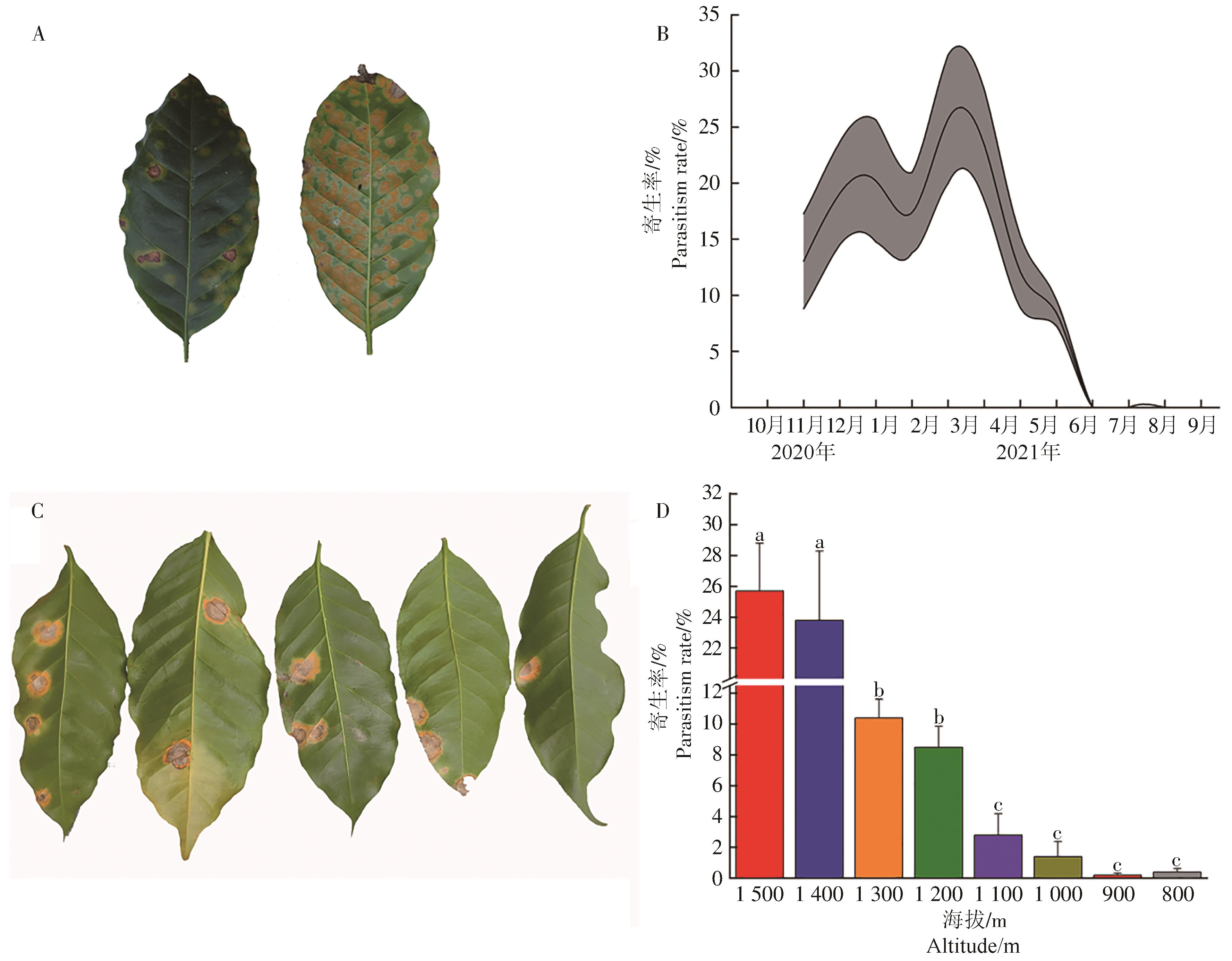
咖啡叶锈病重寄生发生在咖啡叶锈病中后期,无重寄生菌寄生的病斑表面密布黄色或橙黄色锈孢子堆(图4-A),重寄生菌寄生的病斑由中心开始出现灰白色斑块,以同心圆的方式向周围的锈病病斑扩张,咖啡叶锈病病斑开始由中心向边缘褪色,最终黄色锈孢子堆仅剩边缘部位(图4-C)。咖啡叶锈病重寄生从11月至翌年6月发生,具2个高峰,分别为1月和3月,重寄生率分别为(20.19±5.44)%和(25.70±5.72)%,各代峰距约为60 d(图4-B)。海拔梯度间咖啡叶锈病的重寄生率有差异(F=25.585,P<0.01),重寄生率随着海拔的增加而增加,海拔1 500 m的重寄生率显著高于1 400 m以下的6个海拔梯度的,但与海拔1 400 m的差异不显著;海拔1 300 m和1 200 m的咖啡叶锈病重寄生率差异不显著,但显著高于海拔1 100~ 800 m的;海拔1 100~800 m间咖啡叶锈病重寄生率差异不显著(图4-D)。

Figure 4. Mycoparasite fungus of coffee leaf rust
-
咖啡叶锈病是危害小粒咖啡的世界性病害[1]。在干热区的咖啡叶锈病田间症状表现与Kumar等[12]的田间症状描述一致。周年动态监测发现,在干热区保山咖啡叶锈病全年均有危害,具3个危害高峰期,以低温(无霜)、干旱的12月危害最严重。海拔作为一个综合性影响因子,会引起光、温度、湿度、水、土壤等综合的变化[13],进而影响植物病害的发生及危害。本次调查发现,在干热区随着海拔梯度的增加咖啡叶锈病危害加重,但不同海拔梯度咖啡叶锈病的周年危害动态不同,低海拔(700~800 m)危害率低于4%,仅12月和翌年2月发生;海拔900~1 400 m全年有2~5个月不发生;海拔1 500 m咖啡叶锈病危害最严重,全年发生,整体上咖啡叶锈病危害百分比随着海拔高度的增加而增加。建议针对不同海拔梯度的咖啡园,应该制定不同的咖啡叶锈病防控策略。在农业生态系统中病原菌H. vastatrix被重寄生真菌寄生的现象非常普遍,已报道的种类包括Calonectria hemileiae[3]、Lecanicillium lecanii[14]及Trichoderma botryosum[15]等多种。本次调查也发现干热区小粒咖啡种植区也具有丰富的重寄生真菌资源,但尚不明确具体的重寄生真菌种类;调查表明干热区重寄生真菌主要发生期为11月至翌年6月,其发生动态与咖啡叶锈病危害具有明显的滞后性。同时,在海拔1 500 m的咖啡叶锈病重寄生率达(25.70±5.72)%,也表明了该重寄生真菌对咖啡叶锈病具有很好的寄生效果。因此,重寄生真菌作为咖啡叶锈病生防菌具有广阔的应用前景,建议进一步加强该方面的研究工作。但目前尚不明确咖啡叶锈病危害与温度、湿度、降雨量等气象因子的具体关系,因此,有待进一步开展相关研究;同时本调查仅开展为期1年的监测,仍需继续加强咖啡叶锈病的周年监测,为干热区咖啡叶锈病的精准预测及防控提供科学依据。
 DownLoad:
DownLoad: