




-
种群是指在一定的时间和空间内同一种物种的所有个体的集合, 是物种生存、发展和进化的基本单元, 是群落和生态系统重要的组成部分[1]。物种种群的结构和数量的动态变化过程是种群学研究的2个主要的方向[2-3], 种群的结构特征可以反映出种群内的个体的龄级、径级、高度级的分布情况[4-5] ,种群的数量动态变化过程则可以揭示种群个体数量在时间和空间上的变化规律[6]。通过种群的静态生命表、种群的个体存活曲线、种群的数量动态指数、种群个体的生存函数分析和种群的时间序列预测分析均是种群研究的常用方法[7-10], 研究种群目前的生存现状以及与环境的相互作用程度[11-12], 还可预测种群的自然更新的能力以及在受到外界干扰情况下的发展态势, 对物种资源的保护、修复和利用具有重要的意义[13-14]。
红树林湿地是陆地向海洋过渡的区域,主要分布在热带,并延伸至亚热带部分地区,是生产力最高的生态系统之一[15],也是较早受全球气候变化影响的生态系统之一[16], 尤其是在赤道附近的印度洋、太平洋和大西洋沿岸分布的红树林。红树林是海岸带地区的重要的生态屏障[17], 同时也是对全球气候变化最敏感的生态系统之一[18]。全球气候变化将显著影响到红树林的生存状况[19-21]及其分布范围[22]。随着日益增强的人为干扰和全球气候变化的加剧,红树林正面临严重的威胁如生境破碎化和丧失、过度开发、环境污染和外来物种入侵等,多地分布的红树林出现逐步衰退甚至消失的情况[22-26]。
红海榄(Rhizophora stylosa)为常绿灌木或小乔木,是我国大陆沿海红树林生态恢复的优选树种[27-28]。目前,有关红海榄的研究多侧重于个体的结构、功能、生理过程、群落的林分结构、种群的生态位、种分布格局等方面的研究[29-35],但对于红海榄种群结构和数量动态变化特征的研究未见报道东寨港是海南岛红树林分布较为集中的区域,红树林种类丰富和群落发育较好,其中,红海榄是东寨港分布较广的树种之一[36]。2022年,海南省红树林研究院在东寨港国家级红树林保护区塔市村附近选择红树林发育较好的区域,建立了一个1 hm2的红树林固定样地,进行红树林湿地的长期监测。本研究以固定样地内的红海榄种群为研究对象,利用数量生态学研究方法,开展红海榄种群的结构与动态变化特征的分析研究,旨在为东寨港国家级红树林保护区和海南岛红树林湿地的保护和生态修复工程提供依据。
-
样地设在海口市东寨港国家级红树林保护区内,该区域内有红海榄(Rhizophora stylosa)、角果木(Ceriops tagal)、秋茄(Kandelia obovata)、白骨壤(Avicennia marina)、桐花树(Aegiceras corniculatum)、海莲(Bruguiera sexangula)等红树植物,生长良好,保存较完整。土壤为泥沙质和淤泥质。研究区域处于热带季风区海洋性气候区,年平均气温介于23.3~23.8℃,出现过的极端高温为38.9℃,出现过的极端低温为2.6℃,年平均日照量大于2 000 lx,年平均降水量达到1 676.4 mm,雨季始于5月上旬,止于10月下旬。潮汐类型为不规律的半日潮,平均的潮差达到 1.1 m,较适合红树林植物的生长。
-
2021年3月在东寨港塔市附近的红树林区域,沿着潮水方向设置1个100 m×100 m的固定样地(E110°32′13.27861″,N20°0′7.05713″)。按照固定样地建设方法,样地划分为100个10 m×10 m的样方。以样地的西南角为原点,对样地进行调查。记录固定样地内的胸径大于1 cm 的植株的株高、胸径、冠幅及生长状况。
-
采用生态学中径级代替龄级方法,以胸径级确定龄级,对群落中的红海榄进行种群年龄结构分析。本研究根据红海榄的生长特点和野外调查数据,以胸径(Diameter at breast height,DBH)2 cm为标准划分龄级,共划分为10个龄级,其中,胸径 <2 cm的个体划分为I龄级,胸径≥18 cm的个体划分为X龄级(表1)。
表 1 红海榄种群龄级划分
龄级 I II III IV V VI VII VIII IX X 胸径/cm 0<
DBH <22≤
DBH <44≤
DBH < 66 ≤
DBH < 88≤
DBH < 1010 ≤
DBH < 1212 ≤
DBH < 1414 ≤
DBH <1616 ≤
DBH <1818≤
DBH <∞ -
通过计算种群内相邻龄级间个体数量动态变化指数(Vn)和整个种群的个体数量动态变化指数(Vpi),分析海南东寨港保护区内红海榄种群各龄级间和整个种群的数量变化动态特征[37]。由于种群个体数量必然会受到随机的外环境的干扰,因此对整体数量动态变化指数(Vpi)进行修正得到修正后的整体数量动态变化指数(
$ {V'_{pi}} $ )。个体数量动态变化指数的公式[37]:$$ {V_n} = \frac{{{S_n} - {S_{n + 1}}}}{{{\text{max}}\left( {{S_n},{S_{n + 1}}} \right)}} \times 100\text{%} \;, $$ $$\begin{split} \\ V_{p i}=\sum_{n=1}^{k-1}\left(S_n \cdot V_n\right) / \sum_{n=1}^{k-1} S_n \;, \end{split} $$ $$ {V'_{pi}} = \frac{{ \displaystyle \sum\nolimits_{n = 1}^{k - 1} {\left( {{S_n} \cdot {V_n}} \right)} }}{{k \cdot {\text{min}}\left( {{S_1},{S_2},{S_3} \cdot \cdot \cdot ,{S_k}} \right) \cdot \displaystyle \sum\nolimits_{n = 1}^{k - 1} {{S_n}} }} \;, $$ 式中,Vn 为种群从 n 到 n+1 级的个体数量变化动态指数,反映种群相邻龄级个体数量间( 种群整个年龄结构) 关系,正值表示增长,负值表示衰退,零表示稳定。Vpi 为整个种群的数量变化动态指数,Sn 为第 n 龄级的个体数量,k 为种群最大龄级。Pmax为整个种群承担的随机干扰风险概率极大值。V´pi 为种群结构动态对随机干扰的敏感性指数,反映种群数量对未来外部干扰存在着“稀释效应”。
-
本研究根据特定时间生命表的编制方法,编制海南东寨港保护区红海榄种群的静态生命表,通过数据匀滑解决静态生命表绘制时,出现个体死亡率为负的问题[37]。采用Hett提出的数学模型对种群存活曲线进行检验[2]。
使用生存率函数Sx、累计死亡率函数Fx、死亡密度函数fx和危险率函数λx,对该区域的红海榄种群进行生存分析[38]。
$$ S_{i}=S_{1}×S_{2}×S_{3}…×S_{i } \;, $$ $$ F_{i}=1-S_{i} \;, $$ $$ f_{(ti)}= \dfrac{{\mathop S\nolimits_{i - 1} - \mathop S\nolimits_i }}{{\mathop h\nolimits_i }} \;, $$ $$ \left.\lambda_{(t i}\right)=\frac{2\left(1-S_i\right)}{\left[h_i\left(1+S_i\right)\right]} \;,$$ 式中:Si为存活率,hi为龄级宽度。
-
使用时间序列分析中的一次移动平均法,对该区域的红海榄种群各龄级个体数量的变化进行预测,其计测方法如下:
$$ M_t^{\left( 1 \right)} = \frac{1}{n}\sum\limits_{k = t - n + 1}^t {{X_k}} \;,$$ 式中:n表示需要预测的时间(本研究为龄级时间),t代表龄级,Xk为k龄级内的个体数量,Mt(1)定义为经过未来n个龄级时间t龄级的种群个体数量[39]。
-
采用Microsoft word 2010和Microsoft Excel 2010软件整理和分析数据和绘图。
-
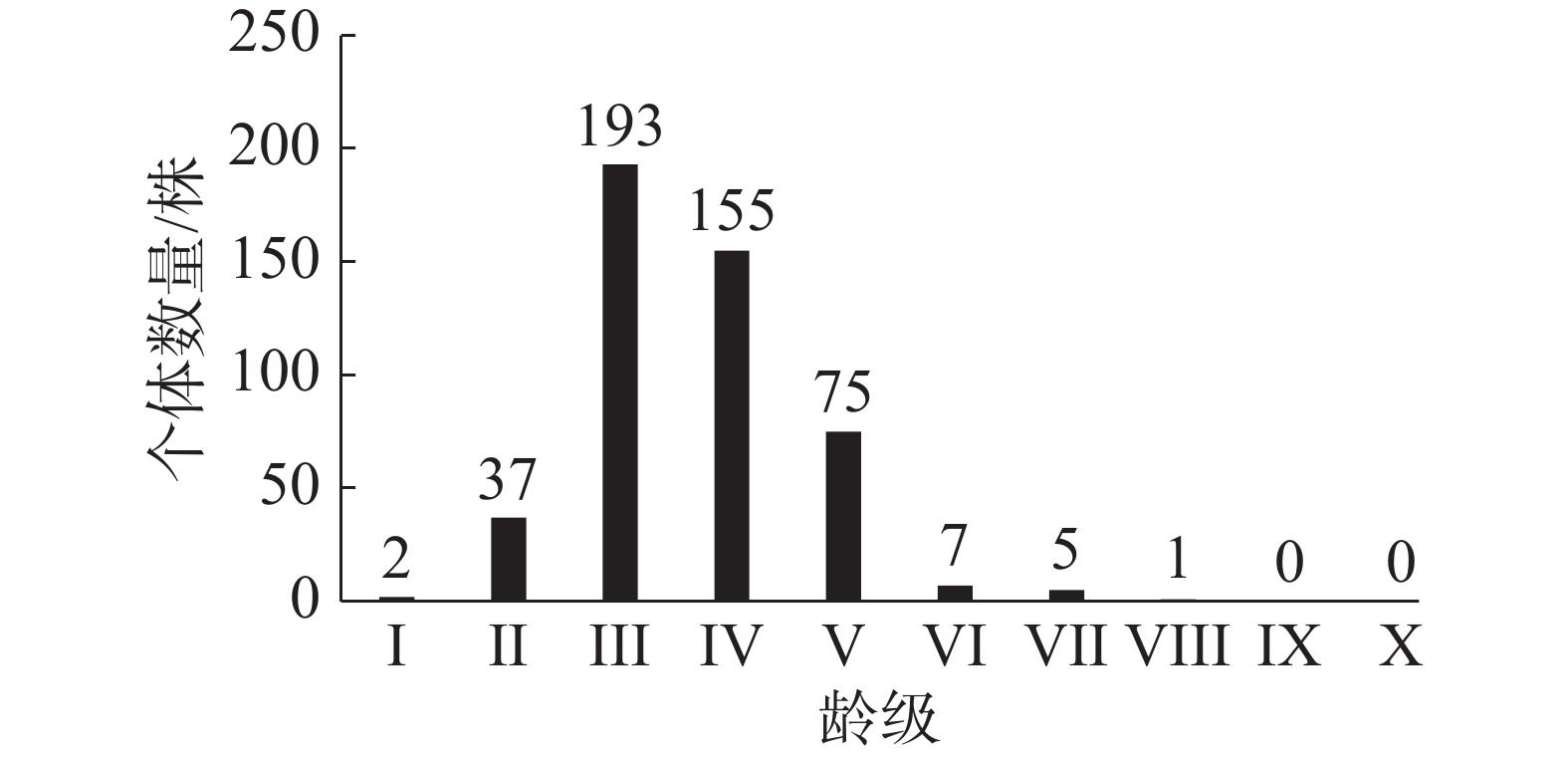
笔者在东寨港固定样地内,调查到红海榄种群个体共475株,其中,个体胸径最大为15.6 cm。从图1可知,该区域红海榄种群的龄级结构为不规则的金字塔类型,III龄级的个体数量相对较多,有193株,占种群个体总数的40.63%。I~III龄级的个体数量之和占种群个体总数的48.84%,IV~V龄级个体数量之和占种群个体总数量的48.43%,而VI~X高龄级的个体数量之和仅占种群个体总数的2.73%。说明低龄级的个体数量较多,III龄级后个体数量开始降低,表明红海榄个体受到环境的影响,种內种间竞争增强,个体数量减少,IV龄级后个体数量急剧降低阶段,可能是由于种內、种间竞争急剧或者受到较强烈外部干扰如病虫害、台风等,引起种群个体死亡增多。
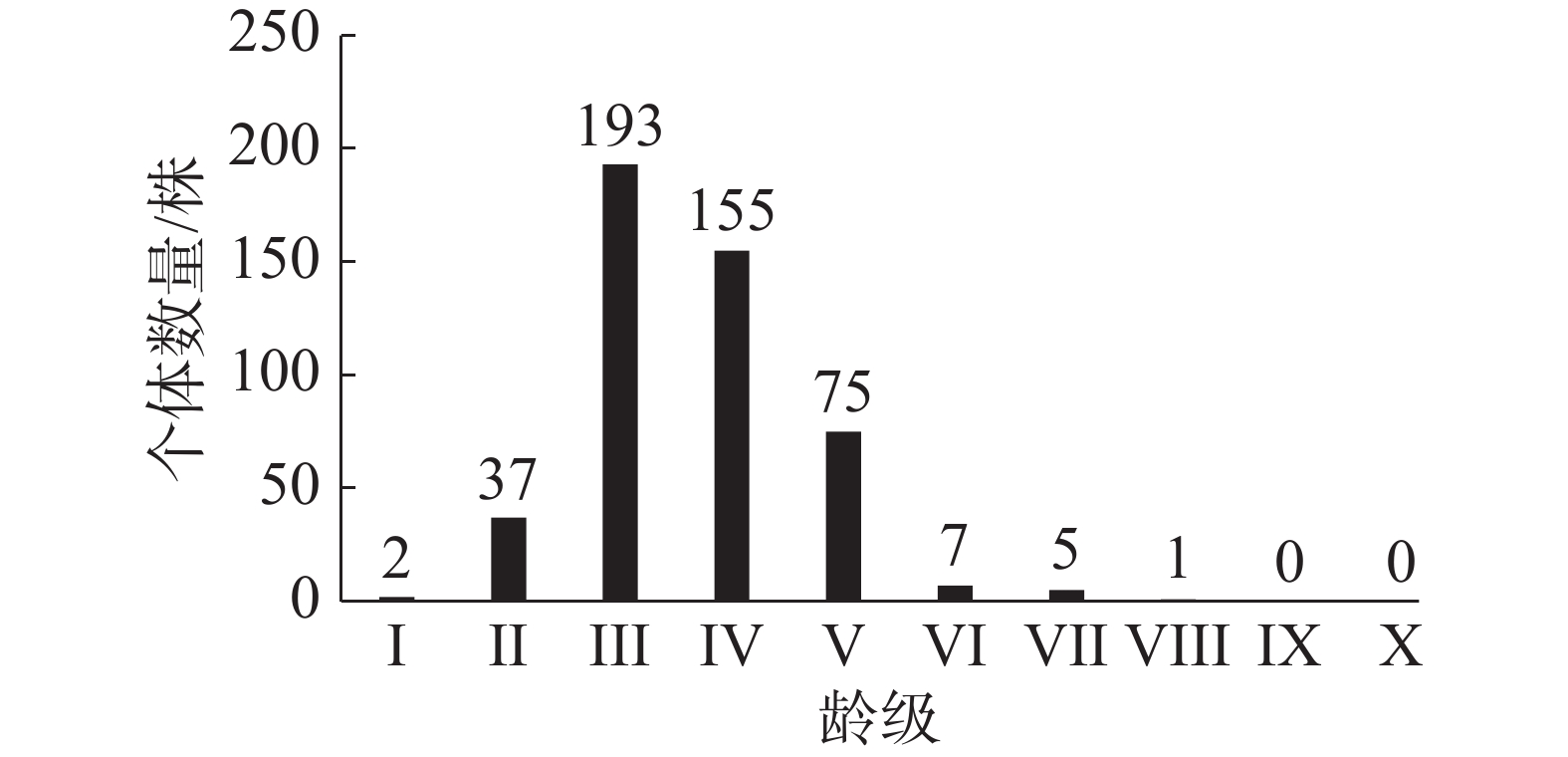
图 1 红海榄种群龄级结构
-
从表2可知,红海榄种群V1~V2小于0,V3~V8大于0。表明红海榄种群整体上表现为“衰退-衰退-增长-增长-增长-增长-增长-增长”的变化趋势。当忽略外环境的干扰时,种群整体的数量变化动态指数Vpi 和V'pi均大于0,说明红海榄种群为增长型种群。但修正后的整个种群的数量变化动态指数V'pi值4.24%,趋近于0,对映出该区域的红海榄种群较不稳定,对外界干扰敏感性较高,受到外界干扰时增长趋势不明显。
表 2 红海榄种群动态变化指数
种群动态指数级 个体数量动态变化指数V(n) Vpi V´pi Pmax 1 2 3 4 5 6 7 8 9 10 指数值/% −94.6 −80.8 19.7 51.6 90.7 28.6 80 100 — — 33.9 4.2 0.13 注:“—”代表空值,由于IX和X龄级的个体数量为0,因此计算后为空值。 -
从表3可知,红海榄不同龄级的个体数量差异明显,I~III龄级的个体数量总和相对较多,表明种群有一定增长能力,但随着龄级的增加,种群个体的标准化存活数量不断降低。生命期望ex可以反映出不同龄级阶段个体的平均生存能力,红海榄种群生命期望值呈现先降后升再降的趋势,其中I~III龄级的生命期望值逐渐降低,说明从I到III龄级各龄级的个体逐步在经历环境的筛选,淘汰个体较多,平均生存能力有所降低。而从IV~VI龄级的生命期望值逐渐增加,说明各龄级个体经过环境的筛选后,个体的适应性增强,平均生存能力有所提升。然而从VI龄级以后,可能是受到生境资源的限制,种內和种间竞争加剧,以及个体进入生理衰老期等因素的造成生命期望值随着龄级的增加逐渐下降。
表 3 红海榄种群静态生命表
龄级 径级 Ax ax lx dx qx Lx Tx lnlx ex Kx Sx Ⅰ 0-2 2 284 1000 225 0.23 887 2335 6.91 2.63 0.26 0.77 Ⅱ 2-4 37 220 775 225 0.29 662 1447 6.65 2.19 0.34 0.71 Ⅲ 4-6 193 156 549 225 0.41 437 785 6.31 1.80 0.53 0.59 Ⅳ 6-8 155 92 324 225 0.70 211 349 5.78 1.65 1.19 0.30 Ⅴ 8-10 75 28 99 67 0.68 65 137 4.59 2.11 1.13 0.32 Ⅵ 10-12 7 9 32 7 0.22 28 72 3.46 2.56 0.25 0.78 Ⅶ 12-14 5 7 25 7 0.29 21 44 3.20 2.08 0.34 0.71 Ⅷ 14-16 1 5 18 7 0.40 14 23 2.87 1.63 0.51 0.60 Ⅸ 16-18 0 3 11 7 0.67 7 9 2.36 1.25 1.10 0.33 Ⅹ 18-∞ 0 1 4 - - 2 - 1.26 - - - 注:Ax :x 龄级存活的个体数量;ax:匀滑后 x 龄级内存活的个体数量; lx: x 龄级的标准化存活个体数量; dx:从 x 到 x+ 1 龄级死亡的个体数量; qx :从 x 到 x+ 1 龄级的种群个体死亡率;Lx :从 x 到 x+1 龄级平均存活的个体数量; Tx :大于等于 x 龄级的存活个体总数; ex:生命期望;lnlx:标准化存活数对数;Kx : 种群消失率; Sx : 种群存活率。 -
从图2可以看出,红海榄种群存活曲线呈现“凹”的变化趋势。通过线性拟合及验证,其存活曲线拟合方程为y=2 708.7e−0.648x,R2为0.97,F值为187.965,P<0.001,这表明该区域的红海榄种群的存活曲线较为符合Deevey-III型曲线,体现出红海榄种群的早期阶段个体死亡率较大,而达到后期阶段个体死亡率趋于平稳的变化趋势。

图 2 红海榄种群存活曲线
-
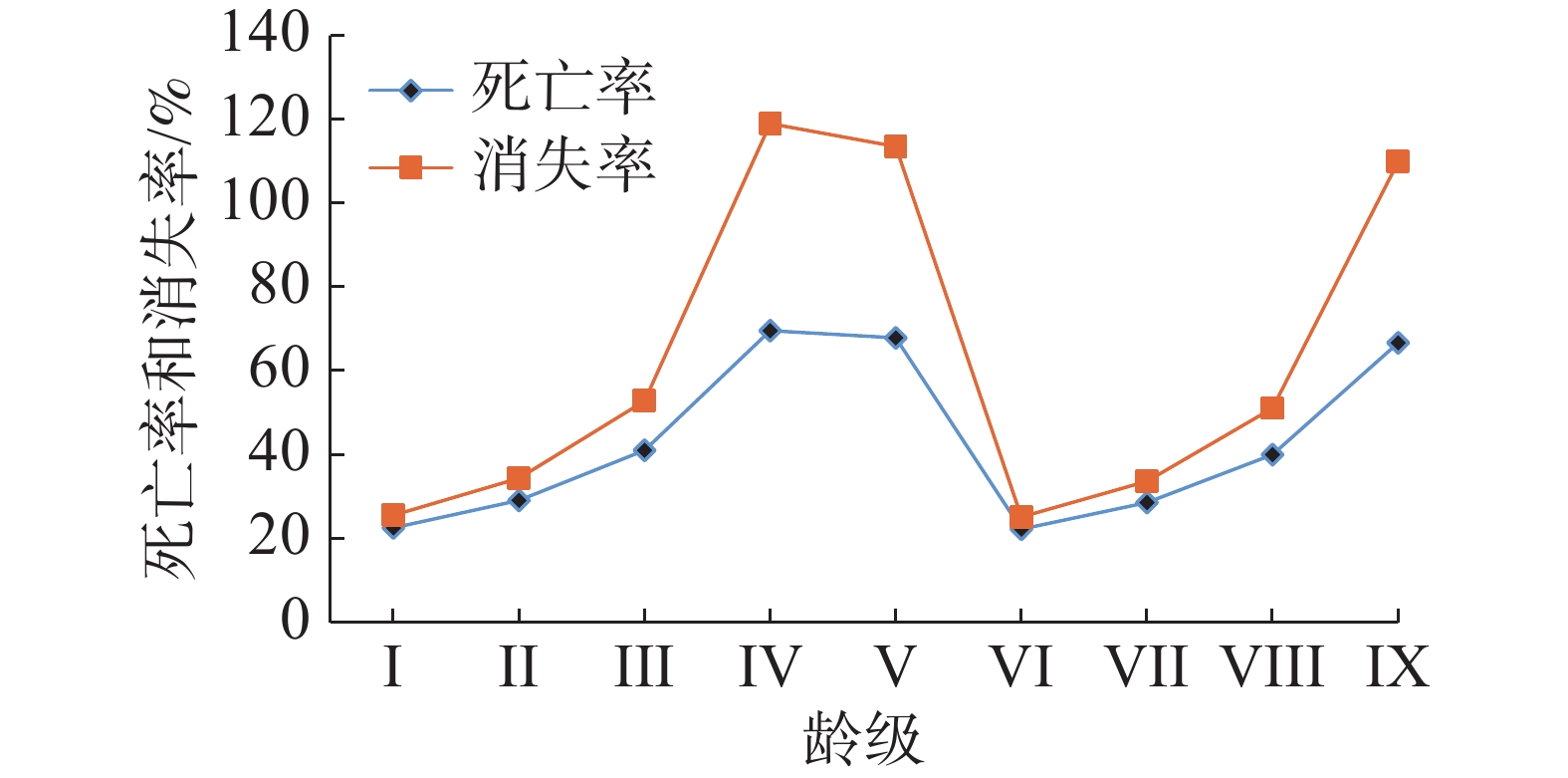
从图3可以看出,该区域红海榄种群的个体死亡率和消失率的变化过程比较相似,龄级增加,死亡率和消失率表现为先增后降的变化趋势。同时结合上述种群数量动态变化的结果可以发现,该区域的红海榄种群数量动态变化较为不稳定。此外,死亡率和消失率均在IV、V和IX龄级出现峰值,可以看出这3个龄级阶段,红海榄种群可能由于外界因素和个体自身的因素导致的种群数量降低。
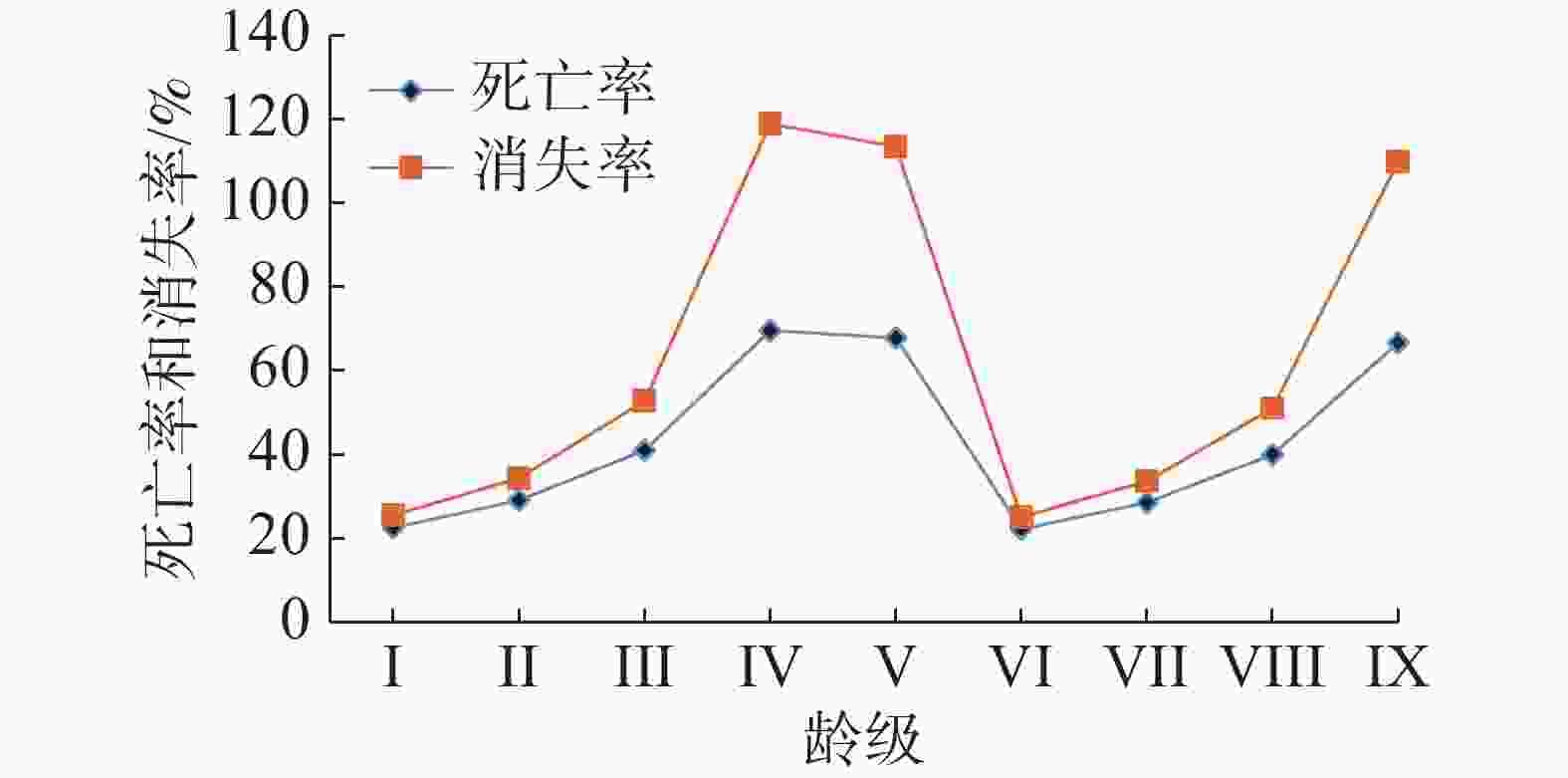
图 3 红海榄种群死亡率与消失率曲线
-
从图4可以看出,随着龄级的增加,红海榄种群的生存率S(i)逐渐降低,而累计死亡率F(i)呈现逐渐上升。其中,在II~III龄级间,两者有交叉点,意味着种群数量在此期间达到平衡,之后累计死亡率超过生存率,种群开始逐渐衰退。在I~V龄级阶段两者变化较为明显,这可能是由于该区域红海榄种群的生存环境条件导致的种群内和种群间强竞争作用造成的。在V龄级以后,两者变化达到极限平稳状态,这表明种群此时的存活压力主要是个体的生理衰老。
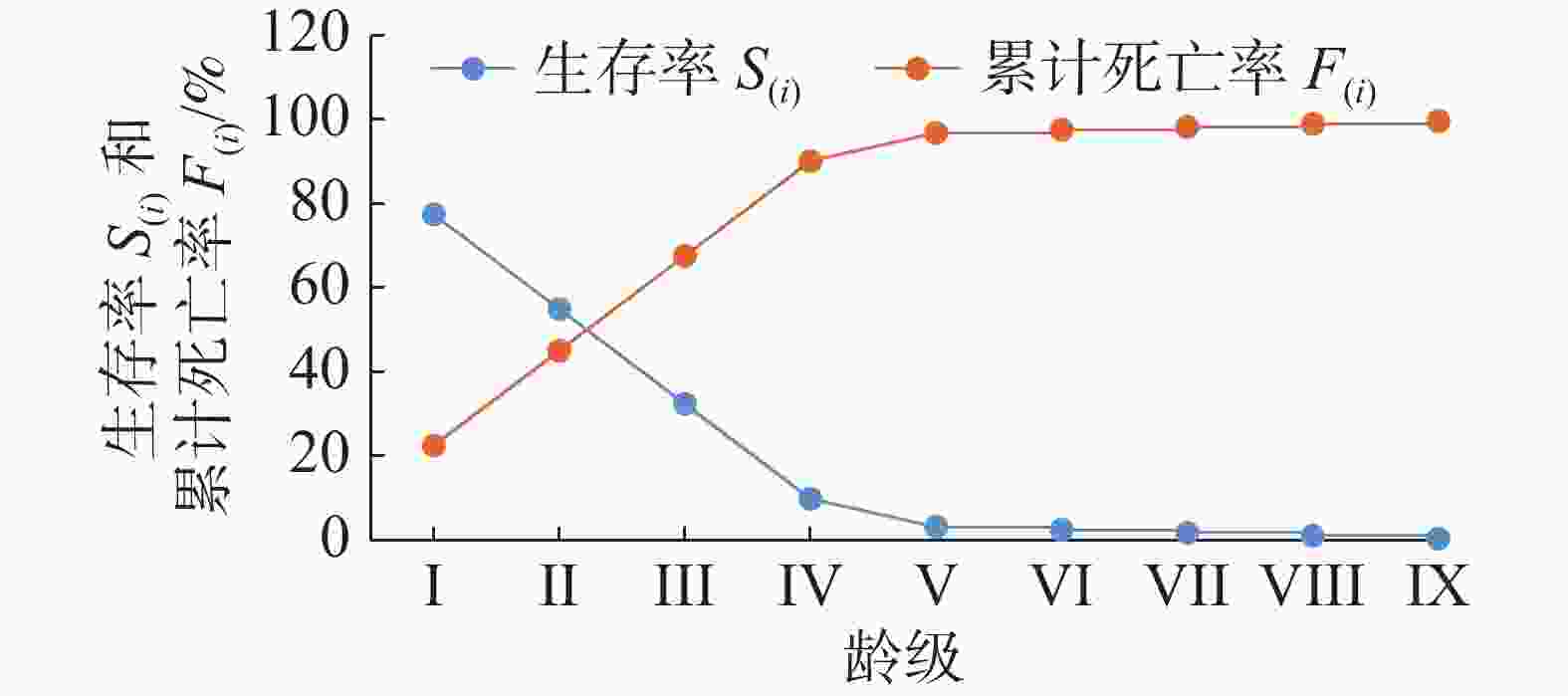
图 4 红海榄种群生存率和累计死亡率曲线
由图5可知,红海榄种群的死亡密度曲线f(ti )呈现递减的变化趋势,在I和II龄级间急剧下降,说明此期间个体数量锐减。在第VI龄级出现最小值(0.001),说明红海榄种群的个体进入了生理衰老期。红海榄种群危险率λ( ti )曲线也呈现递减的变化趋势,在I~II龄级间急剧降低,这反映出,在此阶段红海榄低龄级个体的环境适应性低,面对环境筛选,个体死亡率高,因此危险率较高,随着龄级的增加,种群个体环境适应性逐渐增强,死亡率降低,因此种群的危险率降低。当种群发育到一定的阶段,个体死亡的主要威胁来源于自身的生理寿命,因此,高龄级阶段的危险率几乎趋于平稳的状态。
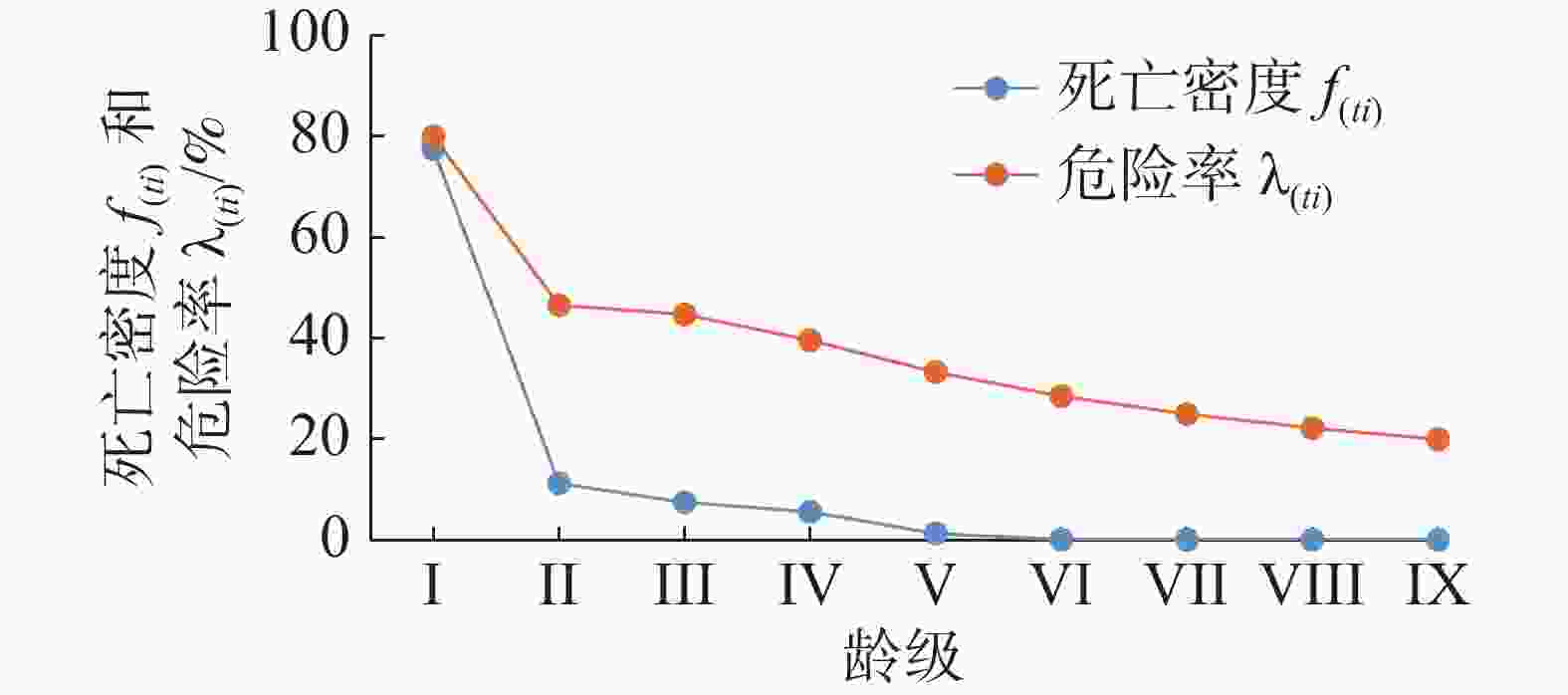
图 5 红海榄种群的死亡密度和危险率曲线
-
根据时间序列分析中的一次平均推移法,对该区域红海榄种群在未来2、3、4、5、6、7、8、9、10个龄级时间后的个体数量变化进行预测。从预测的结果(表4)可以看出,II和III龄级个体数量在经历2~3个龄级时间后种群个体数量均呈现逐渐减少的趋势,表明II和III龄级阶段虽然种群个体数量相对较多,但可以进入高龄级阶段的个体数量却相对较少。IV~X高龄级的个体数量在经历2~10个龄级时间的发育后,种群个体数量均呈现出先增加后减的变化趋势,这说明IV~X龄级的个体具有一定的环境适应性,在一定的龄级时间内存活的个体增多,达到一定的龄级时间后,由于自身原因和外界干扰造成种群数量降低。同时从种群的总个体数量来看,种群随着龄级时间的增加,种群数量逐渐降低。
表 4 红海榄种群数量动态时间序列预测
龄级 各龄级初始个体数量/株 Mt(1) M2(1) M3(1) M4(1) M5(1) M6(1) M7(1) M8(1) M9(1) M10(1) Ⅰ 2 Ⅱ 37 20 Ⅲ 193 115 77 Ⅳ 155 174 128 97 Ⅴ 75 115 141 115 92 Ⅵ 7 41 79 108 93 78 Ⅶ 5 6 29 61 87 79 68 Ⅷ 1 3 4 22 49 73 68 59 Ⅸ 0 1 2 3 18 41 62 59 53 Ⅹ 0 0 0 2 3 15 35 55 53 48 合计 475 474 461 407 342 285 232 173 105 48 注:Mt(1)为经过未来n个龄级时间后各龄级的个体数量。 -
种群的龄级结构分析可以展现个体的发育阶段,同时也可以阐述物种自身生物学特性与生存环境相互作用情况[40]。本研究中红海榄种群的龄级结构呈现不规则的金字塔型,低龄级和高龄级的个体相对较少,中龄级个体数量则相对较多。这与胡刚等对广西北仑河口红树林植物种群研究中桐花树种群的年龄结构较为相似[41]。有研究表明幼龄阶段是种群发育过程中最敏感、最脆弱的阶段[42]。种内和种间竞争以及其他外界环境干扰均会影响植物种群的龄级结构特征[43-45]。本研究红海榄种群的龄级结构的成因,可能是由于本研究区域是以海莲为优势的红树林群落正处于演替的中后期阶段,而红海榄则属于演替的中前期种,因此呈现如此的种群龄级结构特征。此外本研究发现该区域红海榄种群在第Ⅳ龄级后,个体数量锐减,这可能由于该区域曾暴发团水虱危害,红树林植物大量死亡造成的[46]。其他类似的研究均认为种群幼龄阶段个体数量是影响种群发育和恢复的重要的因素[1,6,10]。
-
该区域红海榄种群的幼龄阶段个体较多,意味着该种群具有较好的更新基础,结合种群总体的数量动态变化指数Vpi >V'pi> 0,可以认为该区域的红海榄种群属于增长型的种群,但修正后的总体数量动态变化指数V'pi趋近于0,表明该区域的红海榄种群较为不稳定,易受到外界干扰,种群个体数量增长趋势不显著。宁秋云等[47]在研究广西珍珠湾红树种群结构与动态时也得出类似的结果。由此可以看出,种群数量动态变化指数可以量化的反映种群各龄级阶段个体数量的变化趋势,也能反映种群数量对未来外部干扰存在着“稀释效应”。
种群的静态生命表、存活率、死亡率和消失率不仅可以体现种群的发育状况,还能反映物种与其他物种和生存环境间的相互关系[37,48]。诸多研究者均认为,种群个体的适应能力、生存资源制约的种內和种间竞争加剧,以及个体自身的生理衰老等因素均会导致种群生命期望值降低[49]。本研究发现,该区域红海榄种群的生命期望值呈现先降后升再降的趋势,分别在I和VI龄级出现峰值,低龄级阶段可能是由于个体环境适应力差造成的生命期望的降低,而高龄级的阶段可能是由于生存空间的资源限制引起的种內和种间竞争加剧的结果,较高龄级也可能是由于个体进入生理衰老期导致的生命期望值的降低。胡刚等[41]对广西北仑河口和宁秋云等[47]对广西珍珠湾红树种群的研究均表明,其研究区内不同的红树植物种群的声明期望也各不相同。由此可以看出,不同物种对环境的适应能力的差异,在种群发育过程中生命期望也会有较大的差异。
存活曲线、生存分析函数能够较为直观地反映种群增长与衰退的动态变化过程[50]。本研究发现,该区域的红海榄种群存活曲线呈现“凹”的变化趋势,符合Deevey-III型曲线类型,存活曲线为y=2708.7e−0.648x。早期阶段种群的死亡率较大,后期个体死亡率趋于平稳的变化趋势。研究区内不同的红树植物种群的存活曲线类型均有所差异[41,47]。这就意味着物种及生境条件差异均会影响种群的发育过程。生存分析函数也反映出,该区域的红海榄种群在II和III龄级间,种群个体的生存率曲线和累计死亡率曲线有交叉,意味着种群在该阶段达到平衡,在此之后累计死亡率超过了生存率,种群逐渐衰退。
-
时间序列分析是统计学中的一个重要的分支,其内容非常丰富,在预报分析、林业生产的树木生长、病虫害预测等领域得到广泛应用[51-52]。近年来,被应用到种群研究中进行种群数量的变化预测分析[9,12,37]。本研究通过对红海榄种群数量的时间序列分析发现,第II和III龄级个体数量在经历2~3个龄级时间后种群数量均呈现逐渐下降的趋势,这说明II和III低龄级阶段虽然个体数量较多,但能进入较高的龄级阶段的个体很少。第IV~X龄级阶段的个体在历经2~10个龄级时间发育,种群个体数量先增后降,这说明IV~X龄级的个体具有一定的环境适应性,在一定的龄级时间内存活的个体增多,达到一定的龄级时间后,由于自身原因和外界干扰造成种群数量降低。
从海南东寨港红树林固定样地的红海榄种群的结构及数量动态变化分析结果来看,此区域的红海榄种群为增长型种群,但种群相对不稳定,受到外界干扰后种群个体数量的增长趋势放缓。低龄级的阶段种群的动态变化相对较为剧烈,发展到高龄级阶段种群的动态变化相对平稳。时间序列分析也反映出,虽然低龄级的个体数量较多,经历几个龄级时间后,能发育到较高龄级的个体相对较少,未来的种群将以中高龄级的个体为主体。因此,对于东寨港国家级红树林保护区的红海榄种群应加强长期监测和灾害防治,尤其是加强对低龄级个体的保护和修复。
Population structure and dynamics of Rhizophora stylosa in Dongzhaigang National Mangrove Reserve, Hainan
-
摘要: 为了解东寨港国家级红树林保护区内红海榄(Rhizophora stylosa)种群的动态变化,以保护区内建设的1 hm2的固定样地中的红海榄种群作为研究对象,对其种群的结构与数量动态变化特征进行分析。结果表明:(1)此区域的红海榄种群径级结构为不规则的金字塔型,I~III龄级的个体数量之和占种群个体总数的48.84%,Ⅳ龄级后植株个体数量降低趋势明显。(2)数量动态变化指数反映出该区域的红海榄种群为增长型的种群,但种群较为不稳定,外界干扰下种群数量增长趋势不显著。(3)红海榄种群的存活曲线属于Deevey-Ⅲ类型。(4)红海榄种群的静态生命表、死亡率曲线、消失率曲线以及生存分析反映出该区域的红海榄种群的在低龄级的阶段种群的动态变化相对较为剧烈,发展到高龄级阶段种群的动态变化相对平稳。(5)红海榄种群数量的时间序列分析也反映出,红海榄种群当前虽然低龄级的个体数量较多,但未来几个龄级时间后,能发育到较高龄级的个体相对较少,未来的种群将以中高龄级的个体为主体。东寨港国家级红树林保护区的红海榄种群为增长型种群,但其种群对外界干扰较为敏感,低龄级阶段种群动态变化较为激烈,中、高龄级阶相对较为平稳。因此,应加强对其种群的长期监测以及灾害的防治,尤其要加强对低龄级阶段个体的保护和人工辅助修复。Abstract: It is of great practical significance to carry out relevant research on mangrove plant population to understand the population structure and dynamics of mangroves at the present stage for improving mangrove conservation and ecological restoration. A survey of Rhizophora stylosa population in permanent sample plots covering 1 hm2 in Dongzhaigan National Mangrove Reserve, Haikou, Hainan Province was made to analyze the population structure and dynamics of R. stylosa. The analysis showed that the stem diameter structure of the R. stylosa population in these plots was of irregular pyramid type, with the total number of individuals in age class I to III accounting for 48.84% of the total number of individuals in the population, and the number of individuals in age class Ⅳ being significantly low. The population dynamic change index suggested that the R. stylosa population in this mangrove reserve is expanding, but not stable, and not significantly under external disturbance. The population had a Deevey type II survivorship curve. The static life table, mortality curve, disappearance rate curve and survivorship analysis of the R. stylosa population showed that the population dynamics in this mangrove reserve was relatively drastic at the younger stage, and relatively stable when the population developed into the older stage. The time series analysis of the R. stylosa population reveals that although there are more individuals at the lower age level in the current population, relatively few individuals will develop into the older age level after several age levels in the future, and that the future population will consist of individuals mainly at the middle and older age levels. The population of R. stylosa in Dongzhaigang National Mangrove Reserve is expanding, but is sensitive to external disturbance. The dynamic changes of the population are more intense in the lower age stage, and relatively stable in the middle and old stages. Therefore, the long-term monitoring and disaster prevention of the population should be strengthened, especially the protection and assisted restoration of the individuals at the younger stage.
-
Key words:
- mangrove /
- Rhizophora stylosa /
- population structure /
- population dynamics
-
表 1 红海榄种群龄级划分
龄级 I II III IV V VI VII VIII IX X 胸径/cm 0<
DBH <22≤
DBH <44≤
DBH < 66 ≤
DBH < 88≤
DBH < 1010 ≤
DBH < 1212 ≤
DBH < 1414 ≤
DBH <1616 ≤
DBH <1818≤
DBH <∞ 下载: 导出CSV
下载: 导出CSV
表 2 红海榄种群动态变化指数
种群动态指数级 个体数量动态变化指数V(n) Vpi V´pi Pmax 1 2 3 4 5 6 7 8 9 10 指数值/% −94.6 −80.8 19.7 51.6 90.7 28.6 80 100 — — 33.9 4.2 0.13 注:“—”代表空值,由于IX和X龄级的个体数量为0,因此计算后为空值。
下载: 导出CSV
表 3 红海榄种群静态生命表
龄级 径级 Ax ax lx dx qx Lx Tx lnlx ex Kx Sx Ⅰ 0-2 2 284 1000 225 0.23 887 2335 6.91 2.63 0.26 0.77 Ⅱ 2-4 37 220 775 225 0.29 662 1447 6.65 2.19 0.34 0.71 Ⅲ 4-6 193 156 549 225 0.41 437 785 6.31 1.80 0.53 0.59 Ⅳ 6-8 155 92 324 225 0.70 211 349 5.78 1.65 1.19 0.30 Ⅴ 8-10 75 28 99 67 0.68 65 137 4.59 2.11 1.13 0.32 Ⅵ 10-12 7 9 32 7 0.22 28 72 3.46 2.56 0.25 0.78 Ⅶ 12-14 5 7 25 7 0.29 21 44 3.20 2.08 0.34 0.71 Ⅷ 14-16 1 5 18 7 0.40 14 23 2.87 1.63 0.51 0.60 Ⅸ 16-18 0 3 11 7 0.67 7 9 2.36 1.25 1.10 0.33 Ⅹ 18-∞ 0 1 4 - - 2 - 1.26 - - - 注:Ax :x 龄级存活的个体数量;ax:匀滑后 x 龄级内存活的个体数量; lx: x 龄级的标准化存活个体数量; dx:从 x 到 x+ 1 龄级死亡的个体数量; qx :从 x 到 x+ 1 龄级的种群个体死亡率;Lx :从 x 到 x+1 龄级平均存活的个体数量; Tx :大于等于 x 龄级的存活个体总数; ex:生命期望;lnlx:标准化存活数对数;Kx : 种群消失率; Sx : 种群存活率。
下载: 导出CSV
表 4 红海榄种群数量动态时间序列预测
龄级 各龄级初始个体数量/株 Mt(1) M2(1) M3(1) M4(1) M5(1) M6(1) M7(1) M8(1) M9(1) M10(1) Ⅰ 2 Ⅱ 37 20 Ⅲ 193 115 77 Ⅳ 155 174 128 97 Ⅴ 75 115 141 115 92 Ⅵ 7 41 79 108 93 78 Ⅶ 5 6 29 61 87 79 68 Ⅷ 1 3 4 22 49 73 68 59 Ⅸ 0 1 2 3 18 41 62 59 53 Ⅹ 0 0 0 2 3 15 35 55 53 48 合计 475 474 461 407 342 285 232 173 105 48 注:Mt(1)为经过未来n个龄级时间后各龄级的个体数量。
下载: 导出CSV
-
[1] 赵阳, 刘锦乾, 陈学龙, 等. 洮河上游紫果云杉种群结构特征[J]. 植物生态学报, 2020, 44(3): 266 − 276. doi: 10.17521/cjpe.2019.0077 [2] HETT J M, LOUCKS O L. Age structure models of balsam fir and eastern hemlock [J]. Journal of Ecology, 1976, 64(3): 1029 − 1044. doi: 10.2307/2258822 [3] CHHETRI P K, BISTA R, CAIRNS D M. Population structure and dynamics of Abies spectabilis at treeline ecotone of Barun Valley, Makalu Barun National Park, Nepal [J]. Acta Ecologica Sinica, 2016, 36(4): 269 − 274. doi: 10.1016/j.chnaes.2016.05.003 [4] NUNES Y R F, PETRERE M J. Structure and dynamics of a Cariniana estrellensis (Lecythidaceae) population in a fragment of Atlantic Forest in Minas Gerais, Brazil [J]. Rodriguésia, 2012, 63(2): 257 − 267. [5] 张亚芳, 李登武, 王梅, 等. 黄土高原不同地区杜松种群结构与动态[J]. 林业科学, 2015, 51(2): 1 − 10. [6] 谭一波, 詹潮安, 肖泽鑫, 等. 广东南澳岛中华楠种群结构及动态特征[J]. 生态学杂志, 2010, 29(10): 1901 − 1906. doi: 10.13292/j.1000-4890.2010.0263 [7] HARCOMBE P A. Tree life tables: Simple birth, growth, and death data encapsulate life histories and ecological roles [J]. BioScience, 1987, 37(8): 557 − 568. doi: 10.2307/1310666 [8] 李艳丽, 杨华, 亢新刚, 等. 长白山云冷杉种群结构和动态分析[J]. 北京林业大学学报, 2014, 36(3): 18 − 25. doi: 10.13332/j.cnki.jbfu.2014.03.003 [9] 杨立荣, 张治礼, 云勇, 等. 濒危植物海南龙血树的种群结构与动态[J]. 生态学报, 2018, 38(8): 2802 − 2815. [10] 秦爱丽, 马凡强, 许格希, 等. 珍稀濒危树种峨眉含笑种群结构与动态特征[J]. 生态学报, 2020, 40(13): 4445 − 4454. [11] WANG T, LIANG Y, REN H B, et al. Age structure of Picea schrenkiana forest along an altitudinal gradient in the central Tianshan Mountains, northwestern China [J]. Forest Ecology and Management, 2004, 196(2/3): 267 − 274. doi: 10.1016/j.foreco.2004.02.063 [12] 肖宜安, 何平, 李晓红, 等. 濒危植物长柄双花木自然种群数量动态[J]. 植物生态学报, 2004, 28(2): 252 − 257. doi: 10.17521/cjpe.2004.0037 [13] 陈晓德. 植物种群与群落结构动态量化分析方法研究[J]. 生态学报, 1998, 18(2): 214 − 217. doi: 10.3321/j.issn:1000-0933.1998.02.017 [14] 刘海洋, 金晓玲, 沈守云, 等. 湖南珍稀濒危植物——珙桐种群数量动态[J]. 生态学报, 2012, 32(24): 7738 − 7746. [15] 王友绍. 红树林分子生态学[M]. 北京: 科学出版社, 2019. [16] GILMAN E L, ELLISON J, DUKE N C, et al. Threats to mangroves from climate change and adaptation options: A review [J]. Aquatic Botany, 2008, 89(2): 237 − 250. doi: 10.1016/j.aquabot.2007.12.009 [17] TEMMERMAN S, MEIRE P, BOUMA T J, et al. Ecosystem-based coastal defence in the face of global change [J]. Nature, 2013, 504(7478): 79 − 83. doi: 10.1038/nature12859 [18] LOARIE S R, DUFFY P B, HAMILTON H, et al. The velocity of climate change [J]. Nature, 2009, 462(7276): 1052 − 1055. doi: 10.1038/nature08649 [19] ALONGI D M. Mangrove forests: resilience, protection from tsunamis, and responses to global climate change [J]. Estuarine, Coastal and Shelf Science, 2008, 76(1): 1 − 13. doi: 10.1016/j.ecss.2007.08.024 [20] ALONGI D M. The impact of climate change on mangrove forests [J]. Current Climate Change Reports, 2015(1): 30 − 39. [21] TURNER W R, OPPENHEIMER M, WILCOVE D S. A force to fight global warming [J]. Nature, 2009, 462(7271): 278 − 279. doi: 10.1038/462278a [22] RECORD S, CHARNEY N D, ZAKARIA R M, et al. Projecting global mangrove species and community distributions under climate change [J]. Ecosphere, 2013, 4(3): 1 − 23. [23] 张忠华, 胡刚, 梁士楚. 广西红树林资源与保护[J]. 海洋环境科学, 2007, 26(3): 275 − 279,282. [24] 李春干, 代华兵. 1960—2010 年广西红树林空间分布演变机制[J]. 生态学报, 2015, 35(18): 5992 − 6006. [25] 但新球, 廖宝文, 吴照柏, 等. 中国红树林湿地资源、保护现状和主要威胁[J]. 生态环境学报, 2016, 25(7): 1237 − 1243. doi: 10.16258/j.cnki.1674-5906.2016.07.021 [26] 吴瑞, 陈丹丹, 王道儒, 等. 海南省东寨港红树林资源现状调查分析[J]. 热带农业科学, 2016, 36(9): 62 − 65. [27] 韩维栋, 高秀梅. 雷州半岛红树林生态系统及其保护策略[M]. 广州: 华南理工大学出版社, 2009. [28] 吴钿, 周畅, 刘敏超. 五种海桑属红树植物叶片的结构及其生态适应[J]. 广西植物, 2010, 30(4): 484 − 487. [29] 李润. 红海榄和秋茄次生木质部生态解剖研究[D]. 福州: 福建农林大学, 2018. [30] 周元满, 谢嘉莉, 黄剑坚, 等. 特呈岛红海榄林分结构特征分析[J]. 防护林科技, 2017(12): 5 − 9,37. [31] 邓传远, 郑俊鸣, 张万超, 等. 红海榄木材结构的生态解剖[J]. 植物生态学报, 2015, 39(6): 604 − 615. [32] 陈英华, 严重玲, 李裕红, 等. 盐胁迫下红海榄脯氨酸与活性氧代谢特征研究[J]. 厦门大学学报(自然科学版), 2004, 43(3): 402 − 405. [33] 梁士楚. 红海榄群落演替中种群生态位的研究[J]. 广西科学, 1997, 4(2): 120 − 123,125. [34] 林鹏, 尹毅, 卢昌义. 广西红海榄群落的生物量和生产力[J]. 厦门大学学报(自然科学版), 1992, 31(2): 199 − 202. [35] 周元满, 黄剑坚, 韩维栋, 等. 天然红海榄种群分布格局研究[J]. 广西植物, 2013, 33(4): 496 − 501. doi: 10.3969/j.issn.1000-3142.2013.04.012 [36] 辛欣, 宋希强, 雷金睿, 等. 海南红树林植物资源现状及其保护策略[J]. 热带生物学报, 2016, 7(4): 477 − 483. [37] 张金峰, 葛树森, 梁金花, 等. 长白山阔叶红松林紫椴种群结构与动态特征[J]. 生态学报, 5381 2, 42(13): 5381-5390. [38] 王进, 姚兰, 艾训儒, 等. 鄂西南不同区域亮叶桦种群结构与动态特征[J]. 应用生态学报, 2020, 31(2): 357 − 365. doi: 10.13287/j.1001-9332.202002.010 [39] 王泳腾, 黄治昊, 王俊, 等. 燕山山脉黄檗种群结构与动态特征[J]. 生态学报, 2021, 41(7): 2826 − 2834. [40] SVENSSON J S, JEGLUM J K. Structure and dynamics of an undisturbed old-growth Norway spruce forest on the rising Bothnian coastline [J]. Forest Ecol and Manag, 2001, 151: 67 − 79. doi: 10.1016/S0378-1127(00)00697-6 [41] 胡刚, 黎洁, 覃盈盈, 等. 广西北仑河口红树植物种群结构与动态特征[J]. 生态学报, 2018, 38(9): 3022 − 3034. [42] 张金峰, 程继铭, 闫兴富, 等. 种子特征和播种深度对辽东栎种子萌发和幼苗生长的影响[J]. 广西植物, 2020, 40(2): 226 − 236. doi: 10.11931/guihaia.gxzw201812045 [43] 邱华, 舒皓, 吴兆飞, 等. 长白山阔叶红松林乔木幼苗组成及多度格局的影响因素[J]. 生态学报, 2020, 40(6): 2049 − 2056. [44] 肖翠, 刘帅, 黄珍, 等. 长白山阔叶红松林中影响乔木幼苗存活的关键因素[J]. 生态学报, 2015, 35(19): 6557 − 6565. [45] 刘帅, 侯嫚嫚, 廖嘉星, 等. 生物邻体和生境异质性共同驱动乔木幼苗存活动态[J]. 应用与环境生物学报, 2016, 22(4): 639 − 645. [46] 徐蒂, 廖宝文, 朱宁华, 等. 海南东寨港红树林退化原因初探[J]. 生态科学, 2014, 33(2): 294 − 300. [47] 宁秋云, 赖廷和, 曹庆先, 等. 广西珍珠湾红树种群结构与动态特征[J]. 应用海洋学学报, 2022, 41(1): 42 − 52. doi: 10.3969/J.ISSN.2095-4972.2022.01.006 [48] 张晓鹏, 于立忠, 杨晓燕, 等. 辽东山区天然更新红松幼苗种群结构与动态[J]. 应用生态学报, 2022, 33(2): 289 − 296. [49] 拓锋, 刘贤德, 黄冬柳, 等. 祁连山大野口流域青海云杉种群数量动态[J]. 生态学报, 2021, 41(17): 6871 − 6882. [50] 赖江山, 张谧, 谢宗强. 三峡库区常绿阔叶林优势种群的结构和格局动态[J]. 生态学报, 2006, 26(4): 1073 − 1079. doi: 10.3321/j.issn:1000-0933.2006.04.013 [51] 吴承祯, 洪伟. 林木生长的多维时间序列分析[J]. 应用生态学报, 1999, 10(4): 12 − 15. doi: 10.3321/j.issn:1001-9332.1999.04.004 [52] ZHANG W, ZU Y, YAN X. Comparative study on population age structures between Adenophora lobophylla and A. potaninii [J]. Journal of Forestry Research, 1999, 10(1): 1 − 6. doi: 10.1007/BF02855469 -

 点击查看大图
点击查看大图
图(5) / 表 (4)
计量
- 文章访问数: 379
- HTML全文浏览量: 183
- PDF下载量: 6
- 被引次数: 0









