






-
巴西橡胶树(Hevea brasiliensis Muell. Arg.)是我国重要的热带经济作物,郑冠标等[1]在橡胶树丛枝病研究初报中提出,橡胶树丛枝病在实生苗、芽接苗,幼龄树及已开割的橡胶树上均有发生,针刺病枝有胶乳流出,剖开皮层及木质部未见组织坏死,病害调查发现橡胶树丛枝病发病率在1%~5%之间。陈慕容等[2]通过芽接传病的致病力测定、抗菌素效果反应及电镜观察发现,橡胶树丛枝病是由类菌原体(MLO)和类立克次氏体(RLO)混合侵染所引起的一种传染性病害,且用丛枝病芽片嫁接健康苗木能引起丛枝病和褐皮病,发病率超过20%,结果表明在增殖苗圃中嫁接丛枝病芽片是成龄开割胶树褐皮病的初侵染来源之一。研究证明橡胶树丛枝病病是植原体病害,杨毅等[3]采用分子生物学的相关技术明确了橡胶树丛枝植原体2个株系(RTSF-DS1A与RTSF-DS1B)之间的遗传相关性和亲缘关系。万倩芸[4-5]在植物扁茎形成机制研究中提出扁茎形成的机制主要是顶端分生组织异常和病原菌诱发植物代谢紊乱;在基于转录组学的茅苍术扁茎变异机制研究中结合KEGG分析的研究结果表明,植原体主要是影响植物激素的传导,导致茅苍术发生形态变异。为了探究橡胶树丛枝病发病的原因,笔者运用石蜡切片技术对橡胶树感病后的组织结构变化进行了研究,并运用转录组测序技术,筛选出可能与丛枝病相关的差异基因,进行GO和KEGG分析,旨在探究其功能和代谢通路,找出可能与橡胶树丛枝病组织结构变化相关的通路。
-
供试材料于2019年3月采自海南省儋州市西联苗圃,选择巴西橡胶树(Hevea brasiliensis Muell. Arg.)品系‘热研7-33-97’物候一致的植株,取健康橡胶树叶片和枝条树皮部分及丛枝病橡胶树叶片和枝条树皮部分,用单面刀片切成2组厚度一致的薄片,1组放入FAA固定液(V70%酒精∶V甲醛∶V冰醋酸=90∶5∶5)中保存,用于切片实验;1组放入液氮保存,用于转录组实验。
-
参照蔡海滨的植物石蜡制片改良方法[6],并作部分修改,具体流程如下:采集新鲜材料固定于FAA固定液(V70%酒精∶V甲醛∶V冰醋酸=90∶5∶5)中,抽气,固定。用70%酒精洗涤3次后,用梯度浓度为70%~100%的乙醇彻底脱水(进行碘溴处理),通过梯度性改变正丁醇和酒精比进行透明处理。然后将材料浸蜡(用纯石蜡逐渐代替含正丁醇的石蜡,将正丁醇挥发),浸蜡结束后便立即包埋,将材料从石蜡中取出放入小盒的包埋石蜡中,在蜡未凝固时将材料摆放间隔均匀,进行横切。然后进行修块,将蜡块修整为横切面长为0.5~1 cm 的长方形蜡块,让材料被切面朝上,固定在小的方形木块上,并使蜡块被切两面平形。将材料固定在切片机切片(厚度一般调至5~15 μm),烘片(45 ℃,24 h)后用有机溶剂逐步脱蜡,最后固绿对染,并以中性树胶封存。将制好的枝茎切片和叶片切片放置荧光数码生物显微镜(U_RFL_TOLYMPUS公司)上观察。枝茎切片于4倍镜下观察,病枝茎和健枝茎分别取厚度大、中、小3处测量并计算平均值;叶片切片于40倍镜下观察,病叶与健叶分别取厚度大、中、小3处测量并计算平均值。
-
将样品液氮研磨后,由北京诺禾致源科技股份有限公司进行转录组测序。
-
参照何佳悦基于Kingfisher Flex系统树木叶片RNA的提取方法提取RNA[7],通过Oligo(dT)磁珠富集带有polyA尾的mRNA,随后在NEB Fragmentation Buffer中用二价阳离子将得到的mRNA随机打断,按照NEB普通建库方式进行建库,使用Qubit2.0 Fluorometer进行初步定量,并使用Agilent 2100 bioanalyzer对文库的insert size进行检测,库检合格后进行Illμmina测序。
-
(1)数据质控分析:高通量测序仪测得的图像数据经CASAVA碱基识别转化为序列数据(读数),再经过原始数据过滤、测序错误率检查、GC含量分布检查,获得后续分析使用的clean 读数,将质控后的clean 读数比对到参考基因组上,使用HISAT2比对软件进行比对,根据比对结果,分别统计读数在基因组外显子区域,内含子区域以及基因间区所占的比例。(2)定量分析:采用subread软件中的featureCounts工具对基因进行表达水平的定量分析。(3)差异分析:运用edgeR软件对其表达数据进行统计学分析,筛选样本在不同状态下表达水平差异显著的基因,用火山图进行直观展示。(4)富集分析:采用clusterProfiler软件对差异基因集进行GO功能富集分析,KEGG通路富集分析,GO(Gene Ontology)是描述基因功能的综合性数据库,GO功能富集以P<0.05作为显著性富集的阈值,通过GO富集分析结果,绘制柱状图进行展示。KEGG(Kyoto Encyclopedia of Genes and Genomes)是用于系统分析基因功能、基因组信息和功能信息的数据库,KEGG通路富集以P<0.05作为显著性富集的阈值,从KEGG富集结果中,选取最显著的20个KEGG通路绘制柱状图进行展示,探究其功能及可能参与的调控通路。
-
与健康橡胶树相比,患病橡胶树最明显的表型变化是扁茎、缩节、顶部叶片变小,开割树的丛枝枝条有些会提早干枯;侧芽提前分化和萌发导致丛簇、茎顶端不规则扭曲并木质化且扁枝部位不会生长侧枝和叶(图1,2)。

图 1 健康橡胶树
Figure 1. A healthy rubber tree

图 2 丛枝病橡胶树
Figure 2. A rubber tree infected with witches’ broom disease
-
相比于健康枝茎,患病枝茎周皮、形成层所占比例减小,初生韧皮纤维外侧未见石细胞或石细胞团存在,靠近形成层细胞排列不规则(图3)。枝茎皮组织结构切片测量结果(表1)表明:(1)健康枝茎形成层平均厚度为72.86 μm,占韧皮部比例为9.11%;患病枝茎形成层平均厚度为25.35 μm,占韧皮部比例为3.93%。(2)健康枝茎周皮平均厚度为85.94 μm,占韧皮部比例为10.75%,患病枝茎周皮平均厚度为52.81 μm,占韧皮部比例为8.18%。(3)在次生木质部中,患病枝茎木质部导管平均粗度为122.07 μm,健康枝木质部导管平均粗度为145.17 μm,此差异可能直接影响木质部水分和矿物质元素的运输。对患病枝茎和健康枝茎周皮、韧皮部、形成层3个部位的平均厚度进行测定(图4),方差分析表明,健康枝茎各个部位厚度均值均高于患病枝茎均值,达到显著差异。
表 1 橡胶树健康枝茎与患病枝茎横切组织切片测量结果对比
Table 1. Comparison of cross-sectional tissues of the branches from the healthy and the witches’ broom disease infected rubber trees
结构名称 Structure 平均厚度 Average thickness/μm 患病枝茎 Branches of the infected tree 健康枝茎 Branches of the healthy tree 周皮 Periderm 52.81 85.94 韧皮部 Phloem 645.39 799.81 形成层 Cambium 25.35 72.86 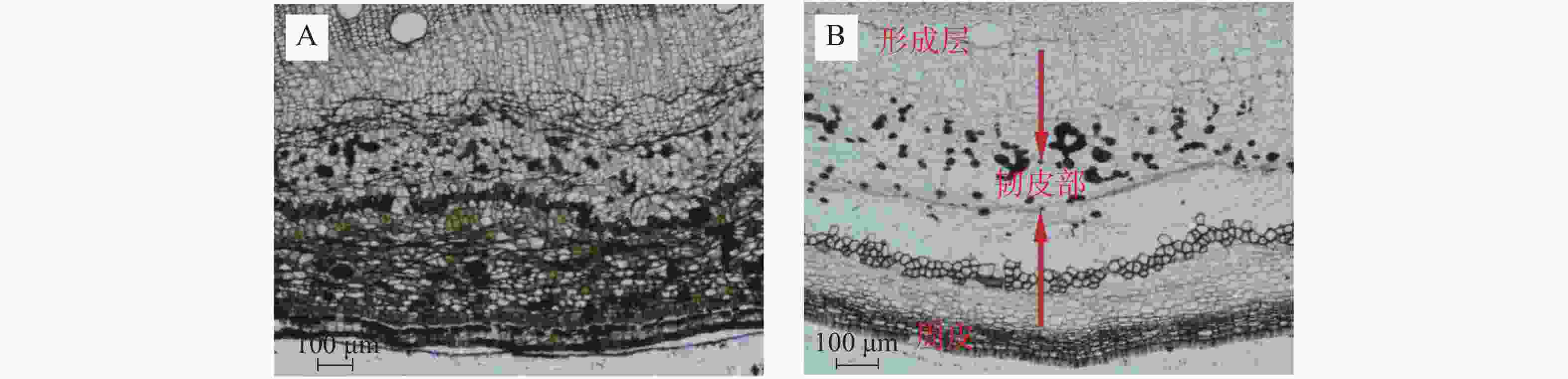
图 3 橡胶树健康枝茎与患病枝茎对比图
Figure 3. Comparison of the branches from the healthy and the witches’ broom infected rubber trees
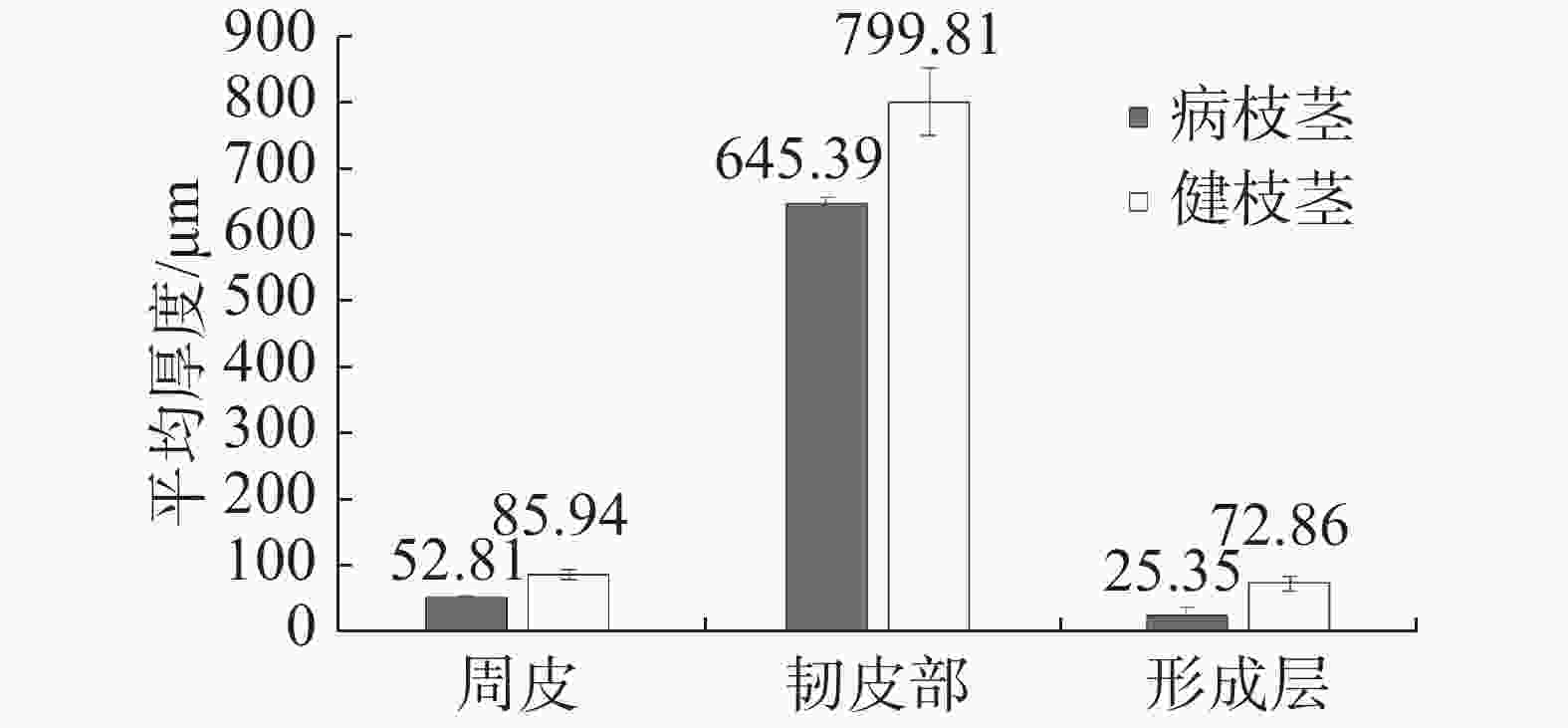
图 4 橡胶树健康枝条与患病枝条枝茎横切组织厚度差异
Figure 4. Analysis of the difference of cross-sectional tissue thickness of the branches from the healthy and witches’ broom infected rubber trees
-
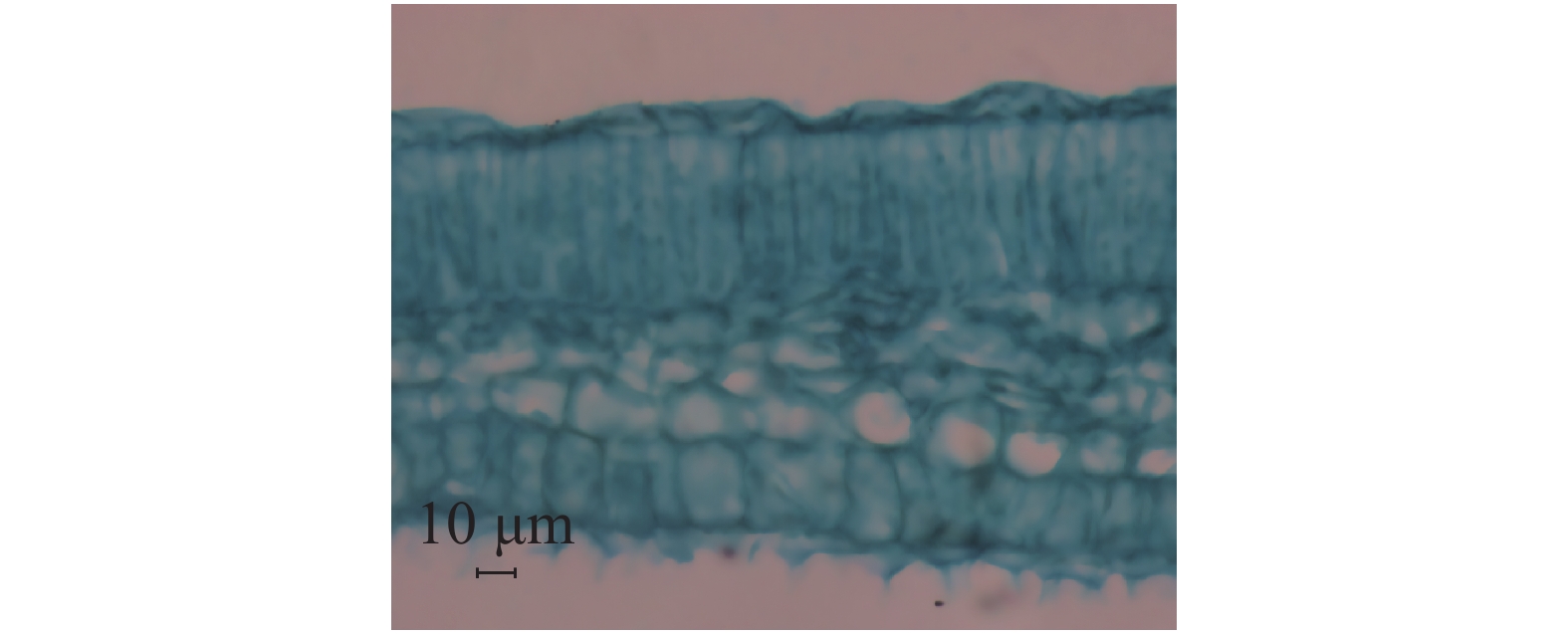
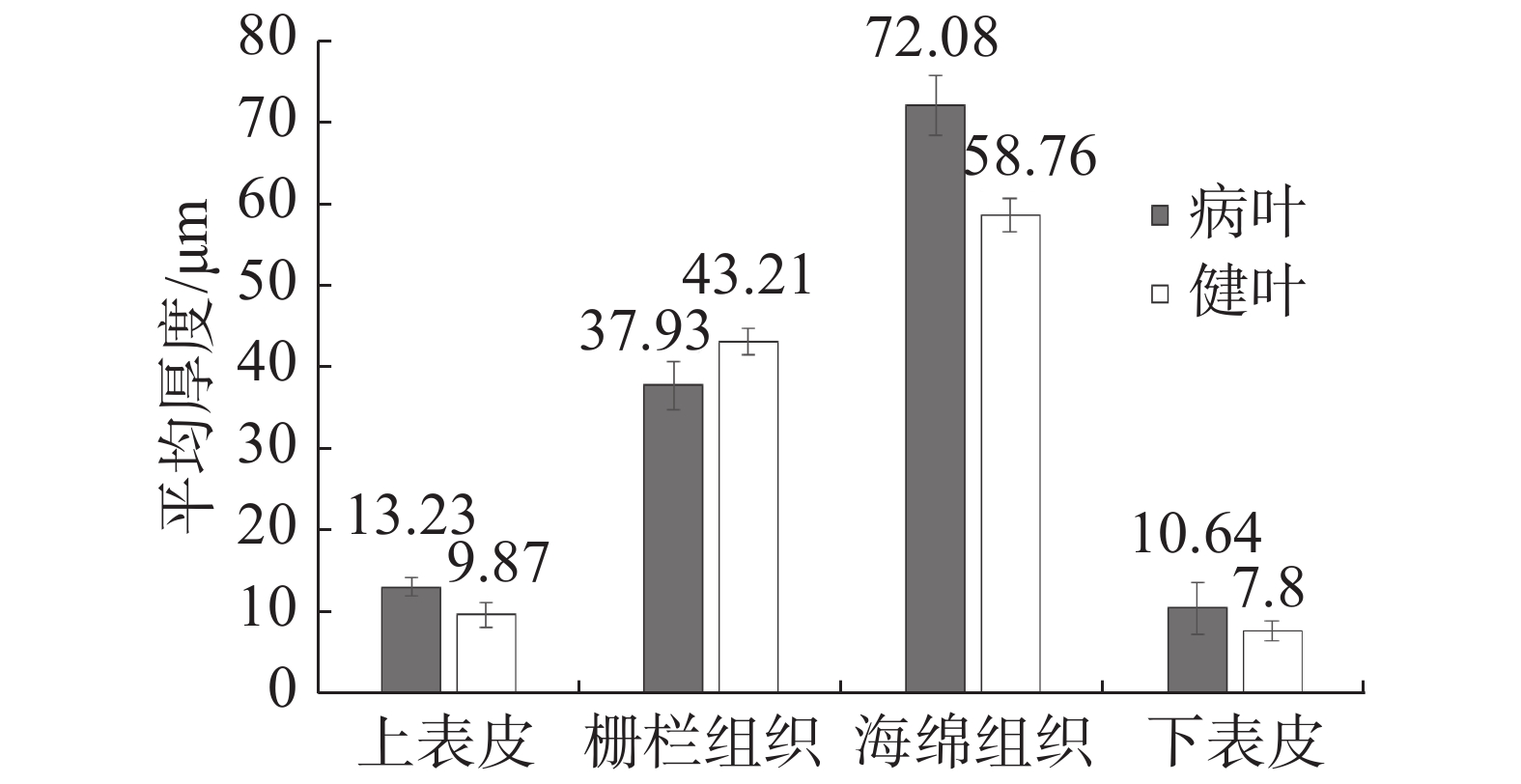
与健康叶片(简称健叶)相比,丛枝病叶片(简称病叶)的栅栏组织变薄,海绵组织变厚且细胞排列不规则,具有海绵组织向栅栏组织过渡的特征;患病叶片叶肉组织细胞排列松散,由长柱形变化为近球形松散海绵组织细胞,细胞间隙明显增大(图5,6)。叶肉组织结构切片测量结果(表2)表明:(1)健叶上表皮平均厚度为9.87 μm,栅栏组织平均厚度为43.21 μm,海绵组织平均厚度为58.76 μm,下表皮平均厚度为7.80 μm。(2)病叶上表皮平均厚度为13.23 μm,栅栏组织平均厚度为37.93 μm,海绵组织平均厚度为72.08 μm,下表皮平均厚度为10.64 μm。(3)石蜡切片显微观察中发现病叶背面毛刺比健叶稀少。对病叶片和健叶片上表皮、栅栏组织、海绵组织、下表皮4个部位的平均厚度进行测定(图7),方差分析结果表明,健叶片栅栏组织厚度均值显著高于病叶片,其他部位厚度均显著低于病枝茎均值,达到显著差异。
表 2 橡胶树健康叶片与丛枝病叶片组织结构厚度对比
Table 2. Comparison of tissue structure thickness of the leaves between the healthy and the witches’ broom disease infected rubber trees
结构名称
Structure平均厚度/μm Average thickness 丛枝病叶片
Leaves of witches’ broom disease infected rubber tree健康叶片
Leaves of the healthy tree上表皮 Upper epidermis 13.23 9.87 栅栏组织 Palisade tissue 37.93 43.21 海绵组织 Spongy tissue 72.08 58.76 下表皮 Lower epidermis 10.64 7.80 总厚度 Total thickness 133.88 119.64 
图 5 橡胶树健康叶片
Figure 5. A leaf blade from the healthy rubber tree

图 6 橡胶树丛枝病叶片
Figure 6. A leaf blade from the rubber tree infected with witches’ broom disease
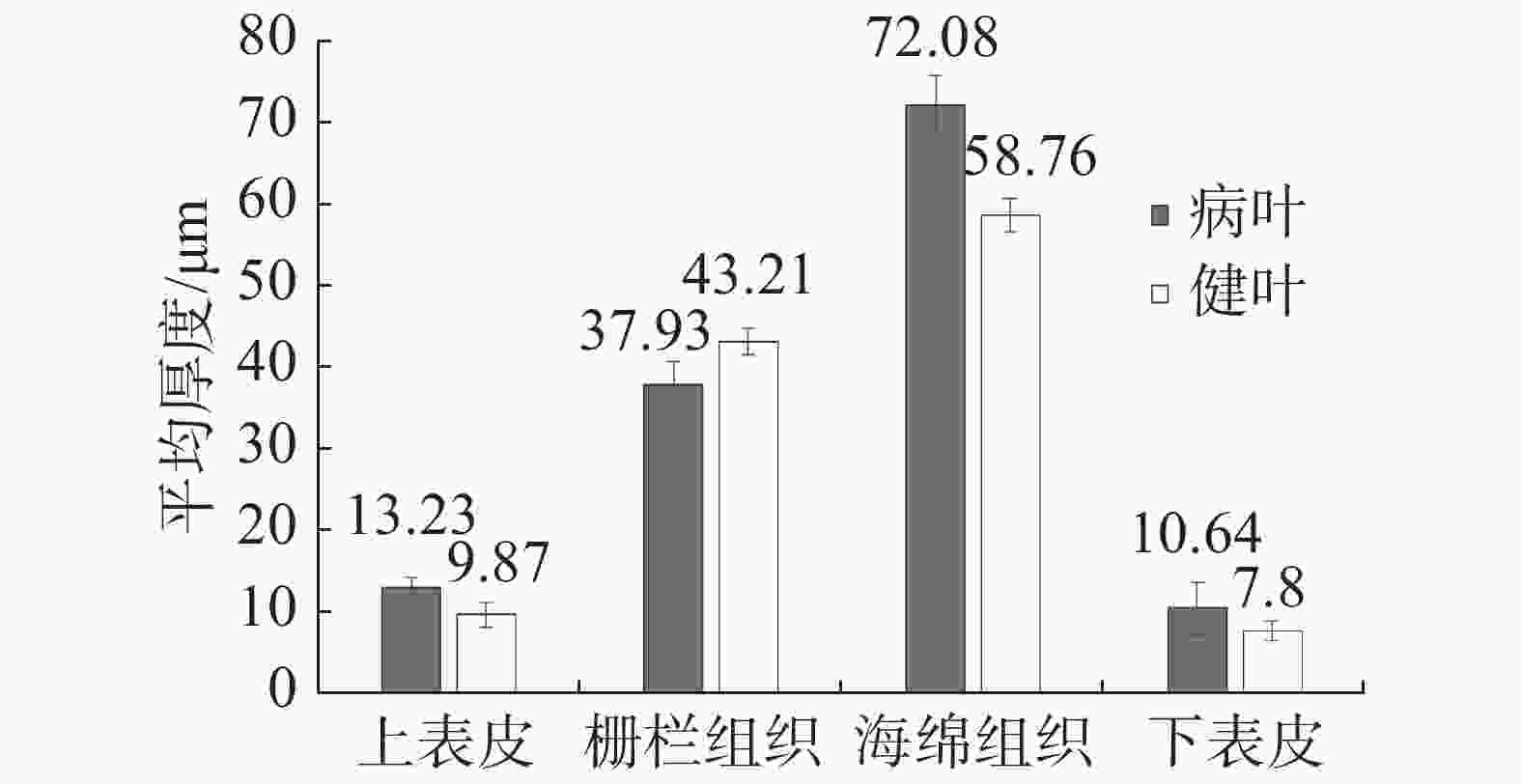
图 7 橡胶树健康叶片与丛枝病叶片组织结构厚度差异性
Figure 7. Analysis of the difference of tissue structure thickness of the leaves between the healthy and the witches’ broom infected rubber trees
-
丛枝病(A)样本总共得到38 902 616个读数,健康(B)样本总共得到43 077 930个读数。丛枝病(A)样本比对率为94.02%,健康(B)样本比对率为93.73%,说明参考基因组组装得较为完善,所测物种与参考基因组一致,且相关实验不存在污染。2个样本的测序读数在基因组区域分布结果(图8)表明:丛枝病(A)样本外显子部分占90.58%,内含子部分所占比例为6.98%,基因间区域占2.45%;健康(B)样本外显子部分占90.21%,内含子部分所占比例为7.49%,基因间区域占2.30%,说明基因注释较为完善。
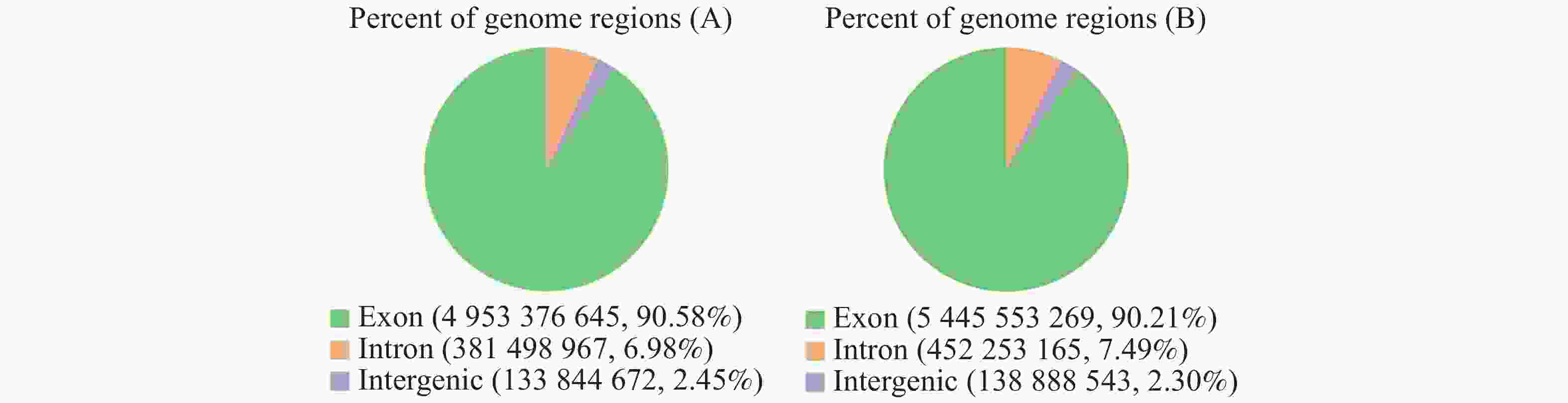
图 8 橡胶树健康样本与丛枝病样本比对区域分布
Figure 8. Comparison of regional distribution of the samples from between the healthy and the witches’ broom disease infected rubber trees
-
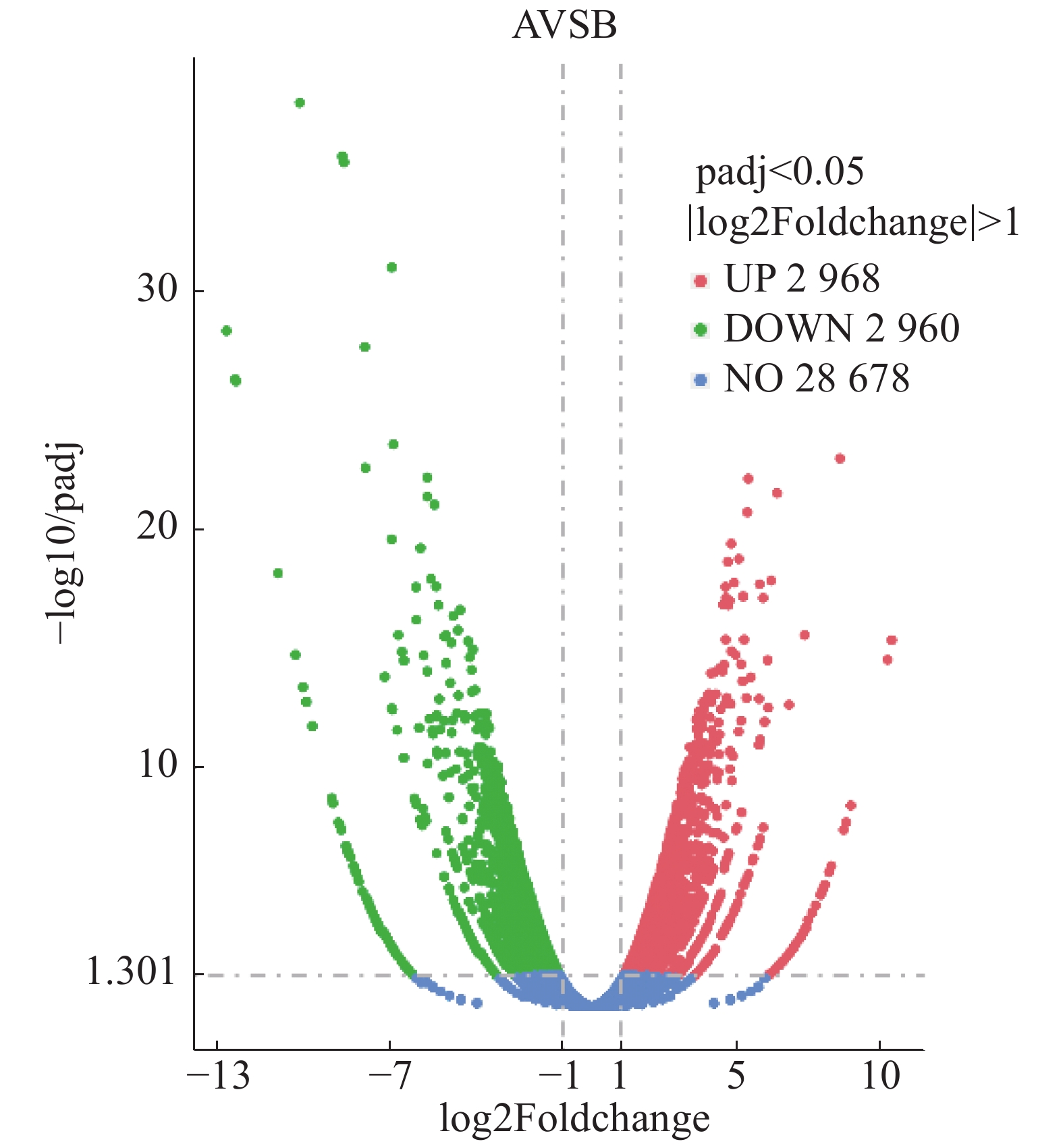
丛枝病(A)样本和健康(B)样本之间有5 928个差异基因,其中上调基因2 968个,下调基因2 960个(图9)。
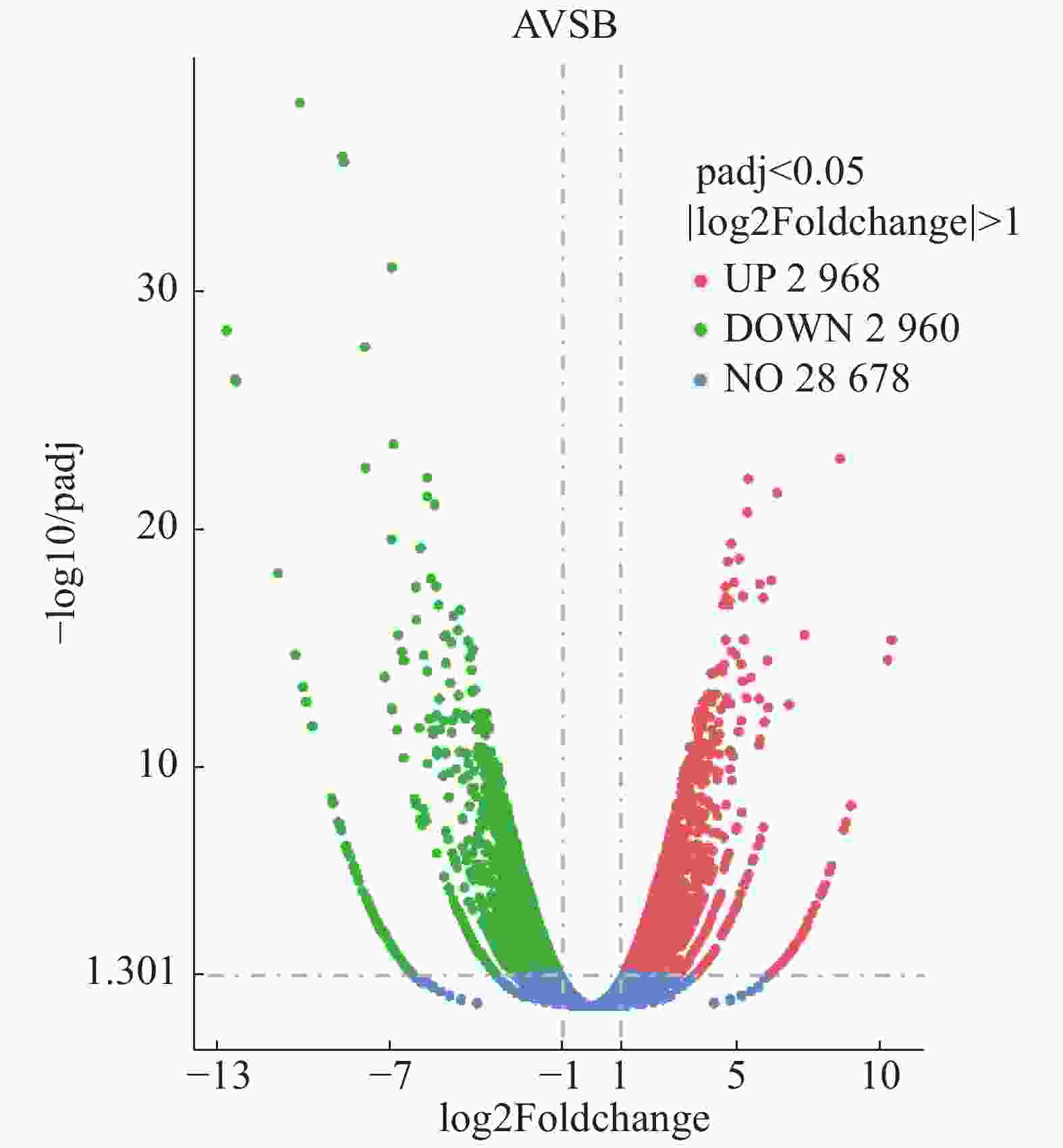
图 9 橡胶树丛枝病样本与健康样本间基因差异表达的火山图
Figure 9. Vocano plot for analysis of gene differential expression of the samples from the witches' broom disease infected and the healthy rubber trees
-
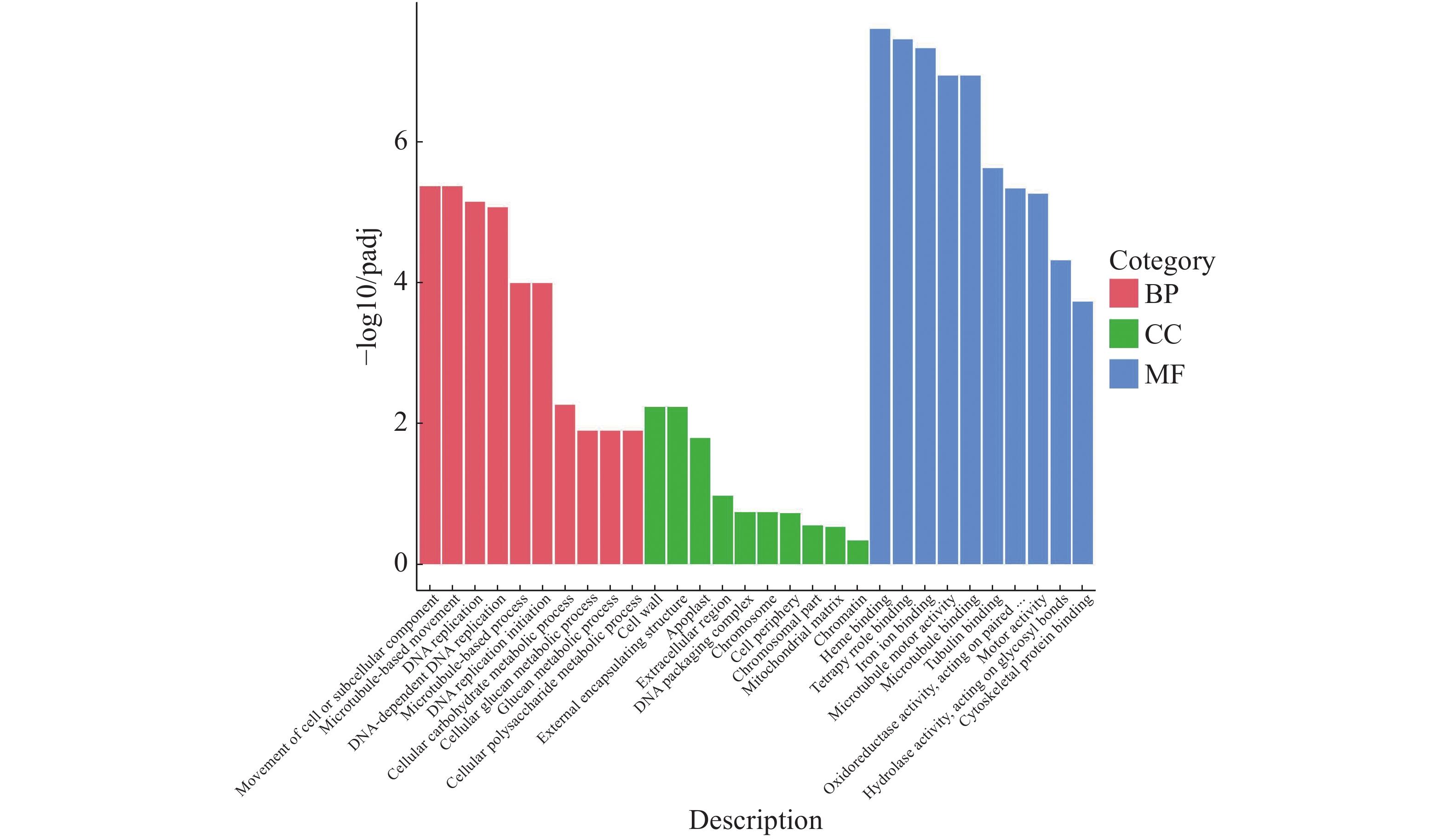
GO富集分析得到的排名前20的生物过程中有2类跟葡聚糖代谢相关,注释到葡聚糖代谢过程的基因有148个(图10)。生物学过程(BP)显著性水平从高到低分别是细胞或亚细胞成分的运动、基于微管的变化、DNA复制、DNA依赖性DNA复制、基于微管的过程、DNA复制启动、细胞碳水化合物代谢过程、细胞葡聚糖代谢过程、葡聚糖代谢过程、细胞多糖代谢过程;细胞成分(CC)显著性水平从高到低分别是细胞壁、外部封装结构、质外体、细胞外区域、DNA包装复杂、染色体、细胞周边、染色体部分、线粒体基质、染色质;分子功能(MF)显著性水平从高到低分别是血红素结合、四吡咯结合、铁离子结合、微管运动结合、微管结合、微管蛋白结合、作用于配对的氧化还原酶活性、运动活动、作用于糖基键的水解酶活性、细胞骨架蛋白结合。
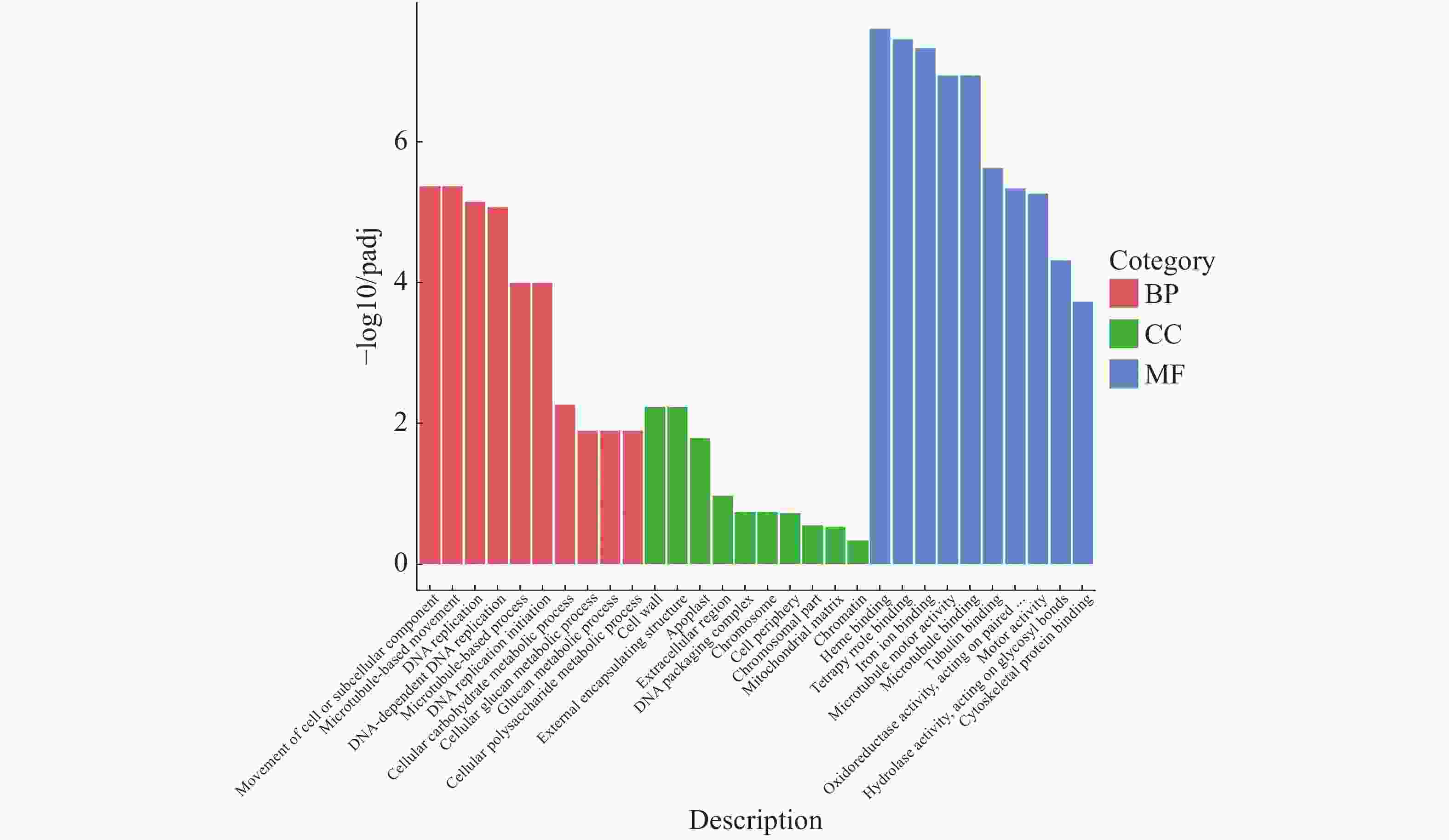
图 10 橡胶树丛枝病样本与健康样本Unigene GO富集性分析
Figure 10. Unigene GO enrichment analysis of the samples from the witches’ broom disease infected and the healthy rubber trees
-
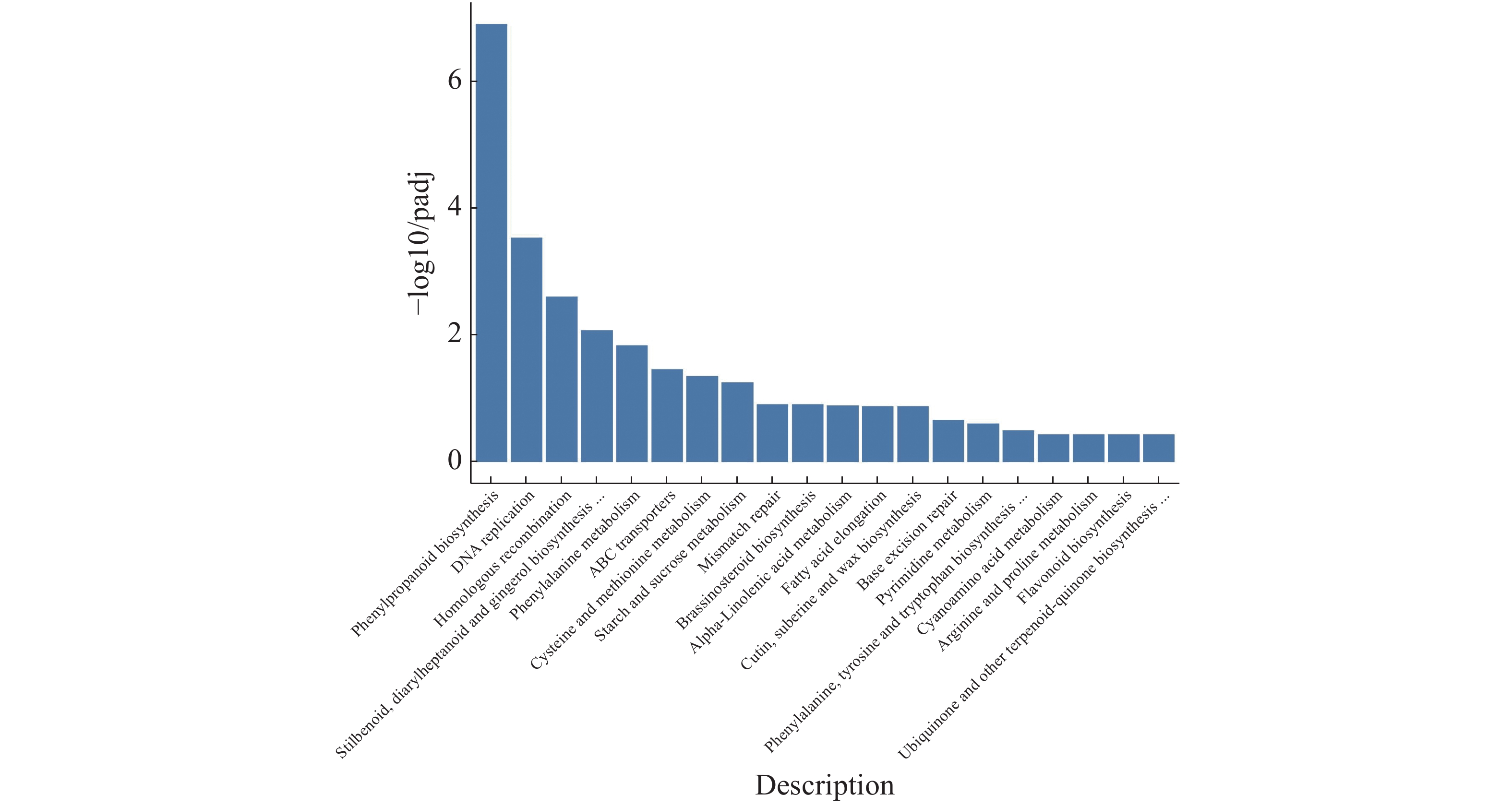
注释到KEGG通路的有527条gene,在最终筛选到的20条关键基因中发现有2条是调控植物黄酮类化合物生物合成的MYB转录因子,油菜素内酯途径也表现为明显下调(图11)。20个最显著通路由高到低依次为:苯丙烷类生物合成、DNA复制、同源重组、stilbenoid,diarylheptanoid和gingerol生物合成、苯丙氨酸代谢、ABC运输车、半胱氨酸和蛋氨酸代谢、淀粉和蔗糖代谢、不匹配修复、油菜素类固醇生物合成、α−亚麻酸代谢、脂肪酸伸长率、角质,小檗碱和蜡生物合成、基础切除修复、嘧啶代谢、苯丙氨酸,酪氨酸和色氨酸生物合成、氰氨基酸代谢、精氨酸和脯氨酸代谢、黄酮类生物合成、泛醌和其他萜类化合物−醌生物合成。
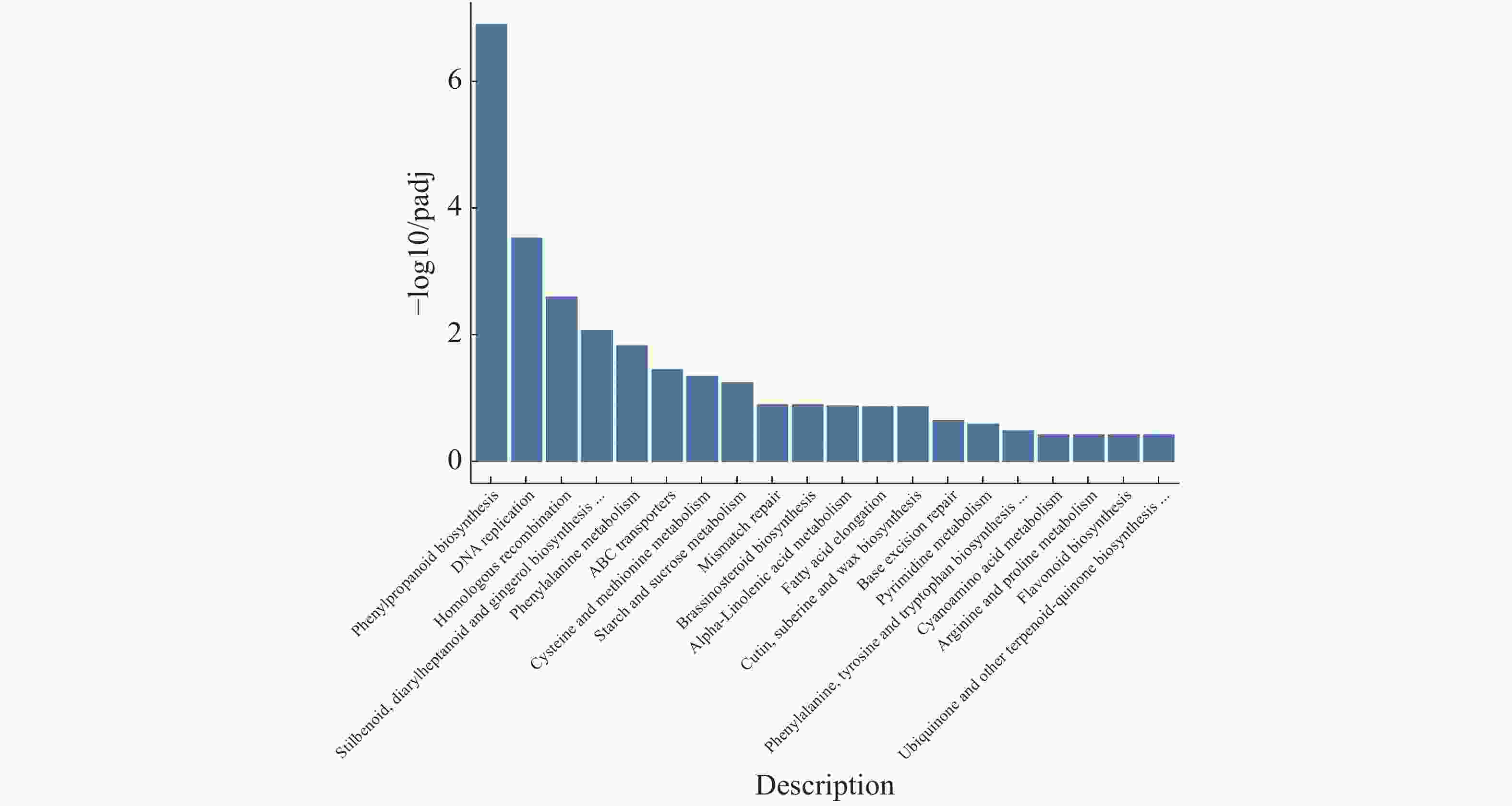
图 11 橡胶树丛枝病样本与健康样本Unigene KEGG富集性分析
Figure 11. Unigene KEGG enrichment analysis of the samples from the witches’ broom disease infected and the healthy rubber trees
-
橡胶树丛枝病的症状为其内部的顶芽变得弱小、侧芽提前分化和萌发导致丛簇、枝干形成层和周皮变薄、木质化程度降低和次生木质部导管缩小,这些结构变化可能导致枝条对水分及矿物质运输能力的减弱和对外部病原的防御变弱,其变化的根本原因也可能是形成层生理生化过程的异常变化。形成层的生理生化变化导致健枝韧皮部容易与木质部剥离,而病枝则不易剥离。病叶相比于健叶,主要是叶背毛刺数目减少、栅栏组织变薄、海绵组织变厚。
由于形成层区域其自身细胞分裂能力对于维管组织的生长非常重要,所以在转录组测序的数据分析中,对患病样本与健康样本差异基因中被注释为细胞分裂以及扩增相关基因进行检索,发现有36个候选基因与细胞分裂有关,与细胞分裂分化相关的基因差异表达,可能是引起丛枝病扁茎的关键因素。
GO富集到148个基因注释到葡聚糖代谢过程,葡聚糖代谢过程在丛枝病发生过程中明显下调。贺新强等[8]在研究杜仲(Eucommia ulmoides )次生木质部分化过程中发现,在形成层及细胞伸展区域,具有葡聚糖的分布,说明葡聚糖在木质部分化和形成层伸展过程中起重要作用。本研究结果表明,在患病橡胶树中,葡聚糖含量减少,形成层区域变薄,分化能力变弱,与文献[8]的结论一致。
KEGG代谢分析,黄酮类化合物和油菜素类固醇生物合成途径表现为显著下调。黄酮类物质广泛存在于植物各器官中,参与花、果实和种子颜色的形成,保护植物抵御紫外线伤害和防止病原微生物侵袭。曹亚兵等[9]在研究泡桐丛枝病发生与代谢组变化的关系中,通过KEGG代谢通路分析表明,有460种代谢物参与111个代谢途径,“黄酮生物合成”是变化最显著的3个代谢途径之一,表明“黄酮生物合成”的变化可能与丛枝病发生有一定关系;曹喜兵[10]在甲基磺酸甲酯对丛枝病泡桐基因表达影响研究中发现黄酮或黄酮类含量随着生物和非生物胁迫的程度而产生变化,黄酮类化合物含量减少增大了病原体入侵的几率,可能与丛枝病的发生有关。已有研究表明植原体侵染植物韧皮部导致丛枝病,曹亚兵等提出“黄酮生物合成”可能与丛枝病发生有一定关系。本实验KEGG结果表明,丛枝病橡胶黄酮类化合物含量减少可能与解剖学中患病橡胶树韧皮部面积小于健康橡胶树有关。
油菜素内酯是一种类固醇类植物激素,与其他植物激素协同作用参与植物的生长发育过程,间接调控植物的光合作用,而且在植物抗逆方面也具有重要的作用。油菜素内酯除了参与到种子萌发、矮化、黑暗中光周期响应、改变气孔分布、延迟开花、雄性不育等发育过程,促进形成层细胞增多,还在细胞伸长和细胞壁扩展及形成层分化中起到重要作用[11-13]。结合本实验转录组KEGG分析得知油菜素内酯在患丛枝病橡胶树体内表现为显著下调,油菜素内酯可能与解剖学中患病叶片栅栏组织和患病茎木质部变化有关。
植原体感染橡胶树导致丛枝病,引起细胞分裂分化相关的基因差异表达,葡聚糖代谢过程、黄酮类生物合成和油菜素类固醇生物合成代谢紊乱,这可能是橡胶树解剖学结构差异的原因。
Tissue Structural Observation and Molecular Mechanism of Witches’ Broom Disease in Rubber Trees
-
摘要: 运用组织结构解剖学方法研究巴西橡胶树(Hevea brasiliensis Muell. Arg.)丛枝病,对比丛枝病橡胶树与健康橡胶树的组织结构差异。采用新一代高通量测序技术Illμmina HiSeq 2000对丛枝病橡胶树和健康橡胶树进行转录组测序。石蜡切片观察结果表明,以健康橡胶枝茎做为对照,丛枝病橡胶枝茎形成层明显变薄,周皮部份占树皮比例明显变小,次生木质部导管变细,初生韧皮纤维外侧没有石细胞存在;丛枝病橡胶叶片栅栏组织较健康叶片变薄,海绵组织较健康叶片变厚。转录组测序结果表明,以健康橡胶树为对照,丛枝病橡胶树的黄酮类化合物和油菜素类固醇生物合成两条代谢途径出现明显紊乱。植原体感染橡胶所引起的代谢紊乱可能是导致橡胶树解剖学结构差异的原因。Abstract: Leaves and branches of a healthy rubber tree (Hevea brasiliensis Muell. Arg.) and a rubber tree infected with witches’ broom disease were sampled for anatomical observation of their tissue structures and for transcriptome sequencing by using a new generation high-throughput sequencing technology Illμmina HiSeq 2000. Paraffin section observation showed that the branches of the rubber tree infected with witches’ broom disease was obviously thinner in cambium, significantly lower in the proportion of the periderm to the bark, thinner in the vessels of the secondary xylem, and absent in stone cells in the outside of the primary phloem fiber as against those of the healthy rubber tree, and that the leaves of the rubber tree infected with the witches’ broom disease was thinner in the palisade tissue and thicker in the spongy tissue as compared to those of the healthy rubber tree. The transcriptome sequencing showed that the two metabolic pathways of the biosynthesis of flavonoids and brassinosteroid in the rubber tree infected with the witches’ broom disease were obviously in disorder as compared to those in the healthy rubber tree. Metabolic disorders caused by phytoplasma –in the rubber tree may be responsible for differences in the anatomical structure of the rubber trees.
-
Key words:
- rubber tree /
- witches’ broom disease /
- tissue structure /
- transcriptome
-
图 3 橡胶树健康枝茎与患病枝茎对比图
A:健康枝茎;B: 患病枝茎。
Fig. 3 Comparison of the branches from the healthy and the witches’ broom infected rubber trees
A: A branch of the healthy tree; B: A branch of infected tree.
图 4 橡胶树健康枝条与患病枝条枝茎横切组织厚度差异
Fig. 4 Analysis of the difference of cross-sectional tissue thickness of the branches from the healthy and witches’ broom infected rubber trees
图 6 橡胶树丛枝病叶片
Fig. 6 A leaf blade from the rubber tree infected with witches’ broom disease
图 7 橡胶树健康叶片与丛枝病叶片组织结构厚度差异性
Fig. 7 Analysis of the difference of tissue structure thickness of the leaves between the healthy and the witches’ broom infected rubber trees
图 8 橡胶树健康样本与丛枝病样本比对区域分布
Fig. 8 Comparison of regional distribution of the samples from between the healthy and the witches’ broom disease infected rubber trees
图 9 橡胶树丛枝病样本与健康样本间基因差异表达的火山图
图中散点代表各个基因,蓝色−圆点表示无显著性差异的基因,红色圆点表示有显著性差异的上调基因,绿色圆点表示有显著性差异的下调基因。横坐标代表基因在丛枝病样本与健康样本中表达倍数变化;纵坐标代表基因表达量变化的统计学显著程度,校正后的p-value越小,log10(校正后的p-value)越大,即差异越显著。
Fig. 9 Vocano plot for analysis of gene differential expression of the samples from the witches' broom disease infected and the healthy rubber trees
In the Figure scattered dots represent genes, blue-dots represent genes with no significant differences, red dots represent up-regulated genes with significant differences, and green dots represent down-regulated genes with significant differences. The abscissa represents the multiple change of gene expression in the samples from the witches' broom disease infected and the healthy rubber trees, and the ordinate represents the statistically significant change of gene expression. The smaller the corrected p-value is, the larger the log10 (corrected p-value) is, that is, the more significant the difference is.
图 10 橡胶树丛枝病样本与健康样本Unigene GO富集性分析
横坐标为GO 3大类下一层级的GO term,纵坐标为注释到该term下的基因个数。3种不同分类表示Go term的3种基本分类(从左往右依次为生物学过程、细胞成分、分子功能)。
Fig. 10 Unigene GO enrichment analysis of the samples from the witches’ broom disease infected and the healthy rubber trees
The abscissa represents GO term, a lower level of the 3 GO major categories, and the ordinate representes the number of genes annotated to the term. The three different categories represent the three basic categories of GO term (The categories from the left to the right are biological processes, cellular components and molecular functions).
图 11 橡胶树丛枝病样本与健康样本Unigene KEGG富集性分析
横坐标为KEGG代谢通路的名称,纵坐标为注释到该通路下的基因个数及其个数占被注释上的基因总数的比例。
Fig. 11 Unigene KEGG enrichment analysis of the samples from the witches’ broom disease infected and the healthy rubber trees
The abscissa represents the name of the KEGG metabolic pathway, and the ordinate represents the proportion of the number of genes annotated to the pathway to the total number of genes annotated.
表 1 橡胶树健康枝茎与患病枝茎横切组织切片测量结果对比
Table 1 Comparison of cross-sectional tissues of the branches from the healthy and the witches’ broom disease infected rubber trees
结构名称 Structure 平均厚度 Average thickness/μm 患病枝茎 Branches of the infected tree 健康枝茎 Branches of the healthy tree 周皮 Periderm 52.81 85.94 韧皮部 Phloem 645.39 799.81 形成层 Cambium 25.35 72.86  下载: 导出CSV
下载: 导出CSV
表 2 橡胶树健康叶片与丛枝病叶片组织结构厚度对比
Table 2 Comparison of tissue structure thickness of the leaves between the healthy and the witches’ broom disease infected rubber trees
结构名称
Structure平均厚度/μm Average thickness 丛枝病叶片
Leaves of witches’ broom disease infected rubber tree健康叶片
Leaves of the healthy tree上表皮 Upper epidermis 13.23 9.87 栅栏组织 Palisade tissue 37.93 43.21 海绵组织 Spongy tissue 72.08 58.76 下表皮 Lower epidermis 10.64 7.80 总厚度 Total thickness 133.88 119.64
下载: 导出CSV
-
[1] 郑冠标, 陈慕容, 陈作义, 等. 橡胶树丛枝病研究初报[J]. 热带作物研究, 1985(1): 19 − 22. [2] 陈慕容, 杨绍华, 郑冠标, 等. 橡胶树丛枝病及其与褐皮病关系的研究[J]. 热带作物学报, 1991(1): 65 − 73. [3] 杨毅, 车海彦, 曹学仁, 等. 橡胶丛枝植原体两个株系的tuf基因克隆及序列分析[J]. 农业科技与信息, 2017(1): 72 − 74. [4] 万倩芸, 陈丽娜, 邓娟, 等. 植物扁茎形成机制研究进展[J]. 分子植物育种, 2017, 15(9): 3717 − 3722. [5] 万倩芸. 基于转录组学的茅苍术扁茎变异机制研究[D]. 武汉: 湖北中医药大学, 2019. [6] 蔡海滨, 涂敏, 胡彦师, 等. 一种优化的橡胶树木质部石蜡切片制作方法[J]. 热带农业科学, 2015, 35(6): 25 − 28. doi: 10.3969/j.issn.1009-2196.2015.06.005 [7] 何佳悦, 王莉, 贾翠蓉, 等. 基于Kingfisher Flex系统树木叶片RNA的提取方法比较及优化[J/OL]. 分子植物育种[2019-11-07][2020-03-03]. http://kns.cnki.net/kcms/detail/46.1068.S.20191107.1307.010.html. [8] 贺新强, 崔克明, 李正理. 杜仲次生木质部分化过程中木质素与半纤维素组分在细胞壁中分布的动态变化[J]. 植物学报, 2001(9): 899 − 904. [9] 曹亚兵, 翟晓巧, 邓敏捷, 等. 泡桐丛枝病发生与代谢组变化的关系[J]. 林业科学, 2017, 53(6): 85 − 93. [10] 曹喜兵. 甲基磺酸甲酯对丛枝病泡桐基因表达影响研究[D]. 郑州: 河南农业大学, 2015. [11] 郭光生. BR对巨桉木质部细胞发育的调控机制研究[D]. 北京: 中国林业科学研究院, 2017. [12] 徐小艳. 油菜素内酯调控杨树维管发育的分子机制分析[D]. 贵阳: 贵州大学, 2019. [13] 闫慧芳, 曾炳山, 范春节, 等. 油菜素内酯对巨桉组培中不定根诱导、苗木生长和茎基部细胞结构的影响[J]. 广西植物, 2016, 36(07): 763 − 767. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 664
- HTML全文浏览量: 104
- PDF下载量: 38
- 被引次数: 0
